Thiolase
Thiolases, also known as acetyl-coenzyme A acetyltransferases (ACAT), are enzymes which convert two units of acetyl-CoA to acetoacetyl CoA in the mevalonate pathway.
| Thiolase, N-terminal domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | Thiolase_N | ||||||||
| Pfam | PF00108 | ||||||||
| InterPro | IPR002155 | ||||||||
| PROSITE | PDOC00092 | ||||||||
| SCOP2 | 1pxt / SCOPe / SUPFAM | ||||||||
| CDD | cd00751 | ||||||||
| |||||||||
| Thiolase, C-terminal domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | Thiolase_C | ||||||||
| Pfam | PF02803 | ||||||||
| InterPro | IPR002155 | ||||||||
| PROSITE | PDOC00092 | ||||||||
| SCOP2 | 1pxt / SCOPe / SUPFAM | ||||||||
| |||||||||
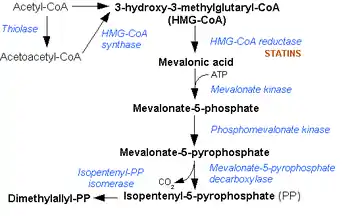
Thiolases are ubiquitous enzymes that have key roles in many vital biochemical pathways, including the beta oxidation pathway of fatty acid degradation and various biosynthetic pathways.[1] Members of the thiolase family can be divided into two broad categories: degradative thiolases (EC 2.3.1.16) and biosynthetic thiolases (EC 2.3.1.9). These two different types of thiolase are found both in eukaryotes and in prokaryotes: acetoacetyl-CoA thiolase (EC:2.3.1.9) and 3-ketoacyl-CoA thiolase (EC:2.3.1.16). 3-ketoacyl-CoA thiolase (also called thiolase I) has a broad chain-length specificity for its substrates and is involved in degradative pathways such as fatty acid beta-oxidation. Acetoacetyl-CoA thiolase (also called thiolase II) is specific for the thiolysis of acetoacetyl-CoA and involved in biosynthetic pathways such as beta-hydroxybutyric acid synthesis or steroid biogenesis.
The formation of a carbon–carbon bond is a key step in the biosynthetic pathways by which fatty acids and polyketide are made. The thiolase superfamily enzymes catalyse the carbon–carbon-bond formation via a thioester-dependent Claisen condensation[2] reaction mechanism.[3]
Function
Thiolases are a family of evolutionarily related enzymes. Two different types of thiolase[4][5][6] are found both in eukaryotes and in prokaryotes: acetoacetyl-CoA thiolase (EC 2.3.1.9) and 3-ketoacyl-CoA thiolase (EC 2.3.1.16). 3-ketoacyl-CoA thiolase (also called thiolase I) has a broad chain-length specificity for its substrates and is involved in degradative pathways such as fatty acid beta-oxidation. Acetoacetyl-CoA thiolase (also called thiolase II) is specific for the thiolysis of acetoacetyl-CoA and involved in biosynthetic pathways such as poly beta-hydroxybutyrate synthesis or steroid biogenesis.
In eukaryotes, there are two forms of 3-ketoacyl-CoA thiolase: one located in the mitochondrion and the other in peroxisomes.
There are two conserved cysteine residues important for thiolase activity. The first located in the N-terminal section of the enzymes are involved in the formation of an acyl-enzyme intermediate; the second located at the C-terminal extremity is the active site base involved in deprotonation in the condensation reaction.
Isozymes
| EC number | Name | Alternate name | Isozymes | Subcellular distribution |
|---|---|---|---|---|
| EC 2.3.1.9 | Acetyl-CoA C-acetyltransferase | thiolase II; Acetoacetyl-CoA thiolase | ACAT1 | mitochondrial |
| ACAT2 | cytosolic | |||
| EC 2.3.1.16 | Acetyl-CoA C-acyltransferase | thiolase I; 3-Ketoacyl-CoA thiolase; β-Ketothiolase | ACAA1 | peroxisomal |
| ACAA2 | mitochondrial | |||
| HADHB | mitochondrial | |||
| EC 2.3.1.154 | Propionyl-CoA C2-trimethyltridecanoyltransferase | 3-Oxopristanoyl-CoA thiolase | ||
| EC 2.3.1.174 | 3-Oxoadipyl-CoA thiolase | β-Ketoadipyl-CoA thiolase | ||
| EC 2.3.1.176 | Propanoyl-CoA C-acyltransferase | Peroxisomal thiolase 2 | SCP2 | peroxisomal/cytosolic |
Mammalian nonspecific lipid-transfer protein (nsL-TP) (also known as sterol carrier protein 2) is a protein which seems to exist in two different forms: a 14 Kd protein (SCP-2) and a larger 58 Kd protein (SCP-x). The former is found in the cytoplasm or the mitochondria and is involved in lipid transport; the latter is found in peroxisomes. The C-terminal part of SCP-x is identical to SCP-2 while the N-terminal portion is evolutionary related to thiolases.[6]
Mechanism
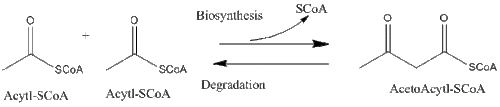
Thioesters are more reactive than oxygen esters and are common intermediates in fatty-acid metabolism.[7] These thioesters are made by conjugating the fatty acid with the free SH group of the pantetheine moiety of either coenzyme A (CoA) or acyl carrier protein (ACP).
All thiolases, whether they are biosynthetic or degradative in vivo, preferentially catalyze the degradation of 3-ketoacyl-CoA to form acetyl-CoA and a shortened acyl-CoA species, but are also capable of catalyzing the reverse Claisen condensation reaction. It is well established from studies on the biosynthetic thiolase from Z. ramigera that the thiolase reaction occurs in two steps and follows ping-pong kinetics.[8] In the first step of both the degradative and biosynthetic reactions, the nucleophilic Cys89 (or its equivalent) attacks the acyl-CoA (or 3-ketoacyl-CoA) substrate,leading to the formation of a covalent acyl-CoA intermediate.[9] In the second step, the addition of CoA (in the degradative reaction) or acetyl-CoA (in the biosynthetic reaction) to the acyl–enzyme intermediate triggers the release of the product from the enzyme.[10] Each of the tetrahedral reaction intermediates that occur during transfer of an acetyl group to and from the nucleophilic cysteine, respectively, have been observed in X-ray crystal structures of biosynthetic thiolase from A. fumigatus.[11]
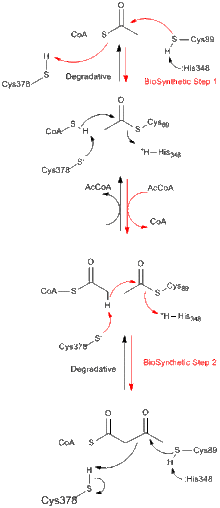
Structure
Most enzymes of the thiolase superfamily are dimers. However, monomers have not been observed. Tetrameters are observed only in the thiolase subfamily and, in these cases, the dimers have dimerized to become tetramers. The crystal structure of the tetrameric biosynthetic thiolase from Zoogloea ramigera has been determined at 2.0 Å resolution. The structure contains a striking and novel ‘cage-like’ tetramerization motif, which allows for some hinge motion of the two tight dimers with respect to each other. The enzyme tetramer is acetylated at Cys89 and has a CoA molecule bound in each of its active-site pockets.[12]
Biological function
In eukaryotic cells, especially in mammalian cells, thiolases exhibit diversity in intracellular localization related to their metabolic functions as well as in substrate specificity. For example, they contribute to fatty-acid β-oxidation in peroxisomes and mitochondria, ketone body metabolism in mitochondria,[13] and the early steps of mevalonate pathway in peroxisomes and cytoplasm.[14] In addition to biochemical investigations, analyses of genetic disorders have made clear the basis of their functions.[15] Genetic studies have also started to disclose the physiological functions of thiolases in the yeast Saccharomyces cerevisiae.[16] Thiolase is of central importance in key enzymatic pathways such as fatty-acid, steroid and polyketide synthesis. The detailed understanding of its structural biology is of great medical relevance, for example, for a better understanding of the diseases caused by genetic deficiencies of these enzymes and for the development of new antibiotics.[17] Harnessing the complicated catalytic versatility of the polyketide synthases for the synthesis of biologically and medically relevant natural products is also an important future perspective of the studies of the enzymes of this superfamily.[18]
Disease relevance
Mitochondrial acetoacetyl-CoA thiolase deficiency, known earlier as β-ketothiolase deficiency,[19] is an inborn error of metabolism involving isoleucine catabolism and ketone body metabolism. The major clinical manifestations of this disorder are intermittent ketoacidosis but the long-term clinical consequences, apparently benign, are not well documented. Mitochondrial acetoacetyl-CoA thiolase deficiency is easily diagnosed by urinary organic acid analysis and can be confirmed by enzymatic analysis of cultured skin fibroblasts or blood leukocytes.[20]
β-Ketothiolase Deficiency has a variable presentation. Most affected patients present between 5 and 24 months of age with symptoms of severe ketoacidosis. Symptoms can be initiated by a dietary protein load, infection or fever. Symptoms progress from vomiting to dehydration and ketoacidosis.[21] Neutropenia and thrombocytopenia may be present, as can moderate hyperammonemia. Blood glucose is typically normal, but can be low or high in acute episodes.[22] Developmental delay may occur, even before the first acute episode, and bilateral striatal necrosis of the basal ganglia has been seen on brain MRI.
References
- Thompson S, Mayerl F, Peoples OP, Masamune S, Sinskey AJ, Walsh CT (July 1989). "Mechanistic studies on beta-ketoacyl thiolase from Zoogloea ramigera: identification of the active-site nucleophile as Cys89, its mutation to Ser89, and kinetic and thermodynamic characterization of wild-type and mutant enzymes". Biochemistry. 28 (14): 5735–42. doi:10.1021/bi00440a006. PMID 2775734.
- Heath RJ, Rock CO (October 2002). "The Claisen condensation in biology". Nat Prod Rep. 19 (5): 581–96. doi:10.1039/b110221b. PMID 12430724.
- Haapalainen AM, Meriläinen G, Wierenga RK (January 2006). "The thiolase superfamily: condensing enzymes with diverse reaction specificities". Trends Biochem. Sci. 31 (1): 64–71. doi:10.1016/j.tibs.2005.11.011. PMID 16356722.
- Baker ME, Billheimer JT, Strauss JF (November 1991). "Similarity between the amino-terminal portion of mammalian 58-kD sterol carrier protein (SCPx) and Escherichia coli acetyl-CoA acyltransferase: evidence for a gene fusion in SCPx". DNA Cell Biol. 10 (9): 695–8. doi:10.1089/dna.1991.10.695. PMID 1755959.
- Yang SY, Yang XY, Healy-Louie G, Schulz H, Elzinga M (June 1990). "Nucleotide sequence of the fadA gene. Primary structure of 3-ketoacyl-coenzyme A thiolase from Escherichia coli and the structural organization of the fadAB operon". J. Biol. Chem. 265 (18): 10424–9. PMID 2191949.
- Igual JC, González-Bosch C, Dopazo J, Pérez-Ortín JE (August 1992). "Phylogenetic analysis of the thiolase family. Implications for the evolutionary origin of peroxisomes". J. Mol. Evol. 35 (2): 147–55. doi:10.1007/BF00183226. PMID 1354266. S2CID 39746646.
- Enzymatic reaction mechanisms. San Francisco: W. H. Freeman. 1979. ISBN 978-0-7167-0070-8.
- Masamune, Satoru; Walsh, Christopher T.; Gamboni, Remo; Thompson, Stuart; Davis, Jeffrey T.; Williams, Simon F.; Peoples, Oliver P.; Sinskey, Anthony J.; Walsh, Christopher T. (1989). "Bio-Claisen condensation catalyzed by thiolase from Zoogloea ramigera. Active site cysteine residues". J. Am. Chem. Soc. 111 (5): 1879, 1991. doi:10.1021/ja00187a053.
- Gilbert HF, Lennox BJ, Mossman CD, Carle WC (July 1981). "The relation of acyl transfer to the overall reaction of thiolase I from porcine heart". J. Biol. Chem. 256 (14): 7371–7. PMID 6114098.
- Mathieu M, Modis Y, Zeelen JP, et al. (October 1997). "The 1.8 A crystal structure of the dimeric peroxisomal 3-ketoacyl-CoA thiolase of Saccharomyces cerevisiae: implications for substrate binding and reaction mechanism". J. Mol. Biol. 273 (3): 714–28. doi:10.1006/jmbi.1997.1331. PMID 9402066.
- Marshall, Andrew C.; Bond, Charles S.; Bruning, John B. (January 25, 2018). "Structure of Aspergillus fumigatus Cytosolic Thiolase: Trapped Tetrahedral Reaction Intermediates and Activation by Monovalent Cations". ACS Catalysis. 8 (3): 1973–1989. doi:10.1021/acscatal.7b02873. hdl:2440/113865.
- Modis Y, Wierenga RK (October 1999). "A biosynthetic thiolase in complex with a reaction intermediate: the crystal structure provides new insights into the catalytic mechanism". Structure. 7 (10): 1279–90. doi:10.1016/S0969-2126(00)80061-1. PMID 10545327.
- Middleton B (April 1973). "The oxoacyl-coenzyme A thiolases of animal tissues". Biochem. J. 132 (4): 717–30. doi:10.1042/bj1320717. PMC 1177647. PMID 4721607.
- Hovik R, Brodal B, Bartlett K, Osmundsen H (June 1991). "Metabolism of acetyl-CoA by isolated peroxisomal fractions: formation of acetate and acetoacetyl-CoA". J. Lipid Res. 32 (6): 993–9. PMID 1682408.
- Middleton B, Bartlett K (March 1983). "The synthesis and characterisation of 2-methylacetoacetyl coenzyme A and its use in the identification of the site of the defect in 2-methylacetoacetic and 2-methyl-3-hydroxybutyric aciduria". Clin. Chim. Acta. 128 (2–3): 291–305. doi:10.1016/0009-8981(83)90329-7. PMID 6133656.
- Kanayama N, Ueda M, Atomi H, Tanaka A (February 1998). "Genetic evaluation of physiological functions of thiolase isoenzymes in the n-alkalane-assimilating yeast Candida tropicalis". J. Bacteriol. 180 (3): 690–8. doi:10.1128/JB.180.3.690-698.1998. PMC 106940. PMID 9457876.
- Price AC, Choi KH, Heath RJ, Li Z, White SW, Rock CO (March 2001). "Inhibition of beta-ketoacyl-acyl carrier protein synthases by thiolactomycin and cerulenin. Structure and mechanism". J. Biol. Chem. 276 (9): 6551–9. doi:10.1074/jbc.M007101200. PMID 11050088.
- Keatinge-Clay AT, Maltby DA, Medzihradszky KF, Khosla C, Stroud RM (September 2004). "An antibiotic factory caught in action". Nat. Struct. Mol. Biol. 11 (9): 888–93. doi:10.1038/nsmb808. PMID 15286722. S2CID 12394083.
- Daum RS, Lamm PH, Mamer OA, Scriver CR (December 1971). "A "new" disorder of isoleucine catabolism". Lancet. 2 (7737): 1289–90. doi:10.1016/S0140-6736(71)90605-2. PMID 4143539.
- Mitchell GA, Fukao T (2001). "Inborn errors of ketone body metabolism". In Scriver CR, Beaudet AL, Sly WS, Valle D (eds.). The metabolic & molecular bases of inherited disease. New York: McGraw-Hill. pp. 2326–2356. ISBN 978-0-07-913035-8.
- Hillman RE, Keating JP (February 1974). "Beta-ketothiolase deficiency as a cause of the "ketotic hyperglycinemia syndrome"". Pediatrics. 53 (2): 221–5. PMID 4812006.
- Robinson BH, Sherwood WG, Taylor J, Balfe JW, Mamer OA (August 1979). "Acetoacetyl CoA thiolase deficiency: a cause of severe ketoacidosis in infancy simulating salicylism". J. Pediatr. 95 (2): 228–33. doi:10.1016/S0022-3476(79)80658-7. PMID 36452.
External links
- Acetyl-CoA+C-Acetyltransferase at the US National Library of Medicine Medical Subject Headings (MeSH)
- Overview of all the structural information available in the PDB for UniProt: P28790 (3-ketoacyl-CoA thiolase) at the PDBe-KB.
- Overview of all the structural information available in the PDB for UniProt: Q56WD9 (3-ketoacyl-CoA thiolase 2, peroxisomal ) at the PDBe-KB.