Oligokyphus
Oligokyphus is an extinct genus of advanced herbivorous cynodonts of the late Triassic to early Jurassic periods. Originally considered to be an early mammal, it is now classified as a Mammaliamorph (nearly a mammal) because Oligokyphus does not have the mammalian jaw attachments and it retains a vestigial joint between the quadrate bone and the squamosal bone in the skull.
| Oligokyphus | |
|---|---|
 | |
| life restoration of Oligokyphus | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Therapsida |
| Clade: | Cynodontia |
| Family: | †Tritylodontidae |
| Genus: | †Oligokyphus Hennig, 1922 |
| Type species | |
| Oligokyphus triserialis Hennig E, 1922 | |
| Species | |
| |
| Synonyms | |
| |
Description
Oligokyphus (meaning "small curved animal"), was a small animal, around 50 centimetres (20 in) in length, belonging to the herbivorous Tritylodontidae family. It resembled a weasel in appearance, with a long and slim body. Oligokyphus was found widely across North America, Europe and China.
Skull and jaw
The teeth of the upper and lower jaw contain bump rows that fit together perfectly in order to maintain an accurate bite. Oligokyphus had a face similar to that of modern mammals, although there were differences in the cheekbones and eye sockets. It had a bony secondary palate and double-rooted cheek teeth. Unlike mammals, the teeth of Oligokyphus did not occlude. The jaw was double jointed, and the neck was flexible, with an atlas and axis and a double occipital condyle.
The teeth were different from those of related cynodonts; there were no canine teeth, and unusually large, rodent-like incisors. There is a large gap, or diastema, separating the cheek teeth from the incisors. The lower jaw of these animals moved back and forth when the mouth was shut so that the food could be chopped up. Oligokyphus had no premaxilla, but did have a lateral extension of the maxilla.
While the postcanines in non-mammalians, such as Oligokyphus, are difficult to differentiate from canines, the lower postcanines of Oligokyphus (also considered to be pre-molars) are defining from other Tritylodonts. On lower postcanine teeth of Trityldonts, two cusps can be found per row; however, Oligokyphus have two rows with three cusps in each row.[1] These cusps, specific to Oligokyphus Tritylodonts, allowed for a well-fitting bite that was particularly good at shredding plant material dense in fiber. The foremost incisors are similar to those of today's rodents, extremely intensified and enlarged. The typical location of canine teeth is left empty with Oligokyphus. Instead, a gap is inserted in this area of the jaw as Oligokyphus lack the teeth commonly known as canines.[2]
Cladistics/Phylogeny
Oligokyphus is in the family Tritylodontidae. The family is named after the shape of their teeth. Tritylodontidae means "three knob teeth". The members of this family were all small to medium-sized advanced synapsids with combined specialized structures for herbivorous eating. The first Tritylodont was found in South Africa in upper Jurassic rocks. It was first thought to be one of the earliest mammals. This classification has since been adjusted. These non-mammals became progressively more mammal-like. They are now classified as the closest relatives to the mammals and this is supported by their high, flat, crested jaw, large zygomatic arches, well developed secondary palate, and specialized dentition.
There have also been comparisons between the cranial nerves of Tritylodonts and mammals. The shoulder girdle and forelimb structures were suggestive of digging animals. These animals were extremely active and burrowed in leaf litter and dirt, which suggests characteristics of rodents and rabbits. They naturally had a metabolism that was partially or completely endothermic. They were thought to be driven out by relatives such as mammals competing for the same ecological niches. Another reason that this animal could have gone extinct was due to new plant development. Some flowering plants, or angiosperms, could have been detrimental to these animals since they may not have been used to eating new plants.
Oligokyphus is placed into the subgroup Probainognathia. This forms a monophyletic group with the tritheledontid Pachygenelus.
Fossil finds
Though Oligokyphus is very widespread, it was not until 1953 that representatives of this group were found. Information was first collected from the Kayenta Formation on Comb Ridge in northeastern Arizona. Numerous specimens of Oligokyphus were obtained by Harvard University and the Museum of Northern Arizona in the "Silty Facies". Many fossils have also been found throughout the UK, Germany and China. Some very small fragment remains have also been found in Antarctica. By these fossil records, one can see that Oligokyphus have a vertical humerus and a minor trochanter. This broad distribution indicates that there were no barriers to separate this terrestrial vertebrate.
Paleoecology
Habitat
Oligokyphus were small tetrapod, terrestrial animals. They have long been considered as mammaliomorphs, a link between earlier synapsids and modern mammals. It is believed these animals were primarily land dwelling, living amongst small shrubs or bushes. It is also thought that Oligokyphus fed on seeds or nuts, as their teeth resemble those of modern animals that also feed on seeds and nuts.[2] It is rather difficult to estimate the social behaviors of Oligokyphus as most of it does not preserve in the fossil record. However, considering the conditions on the planet during the times that Oligokyphus was alive and thriving (late Triassic and early Jurassic) and also the locations of which fossils of these animals were found, some educated predictions can be made about their metabolism and feeding habits. Oligokyphus, with its conveniently placed leg and hip structures, likely was quick-moving and fed off of low-lying plant life. With its long weasel-like body, it may have even been possible for Oligokyphus to reach higher vegetation simply by standing on its hind legs. It probably had good use of its hands to manipulate seeds and other digestively pleasing foods. There has not been any support showing Oligokyphus had the ability to climb vertically, as some rodents are capable of doing today.
A definitive radiometric dating of the area preserved in the formation where Oligokyphus lived has not yet been made, and the available stratigraphic correlation has been based on a combination of radiometric dates from vertebrate fossils, magnetostratigraphy and pollen evidence.[3] It has been surmised that the Kayenta Formation was deposited during the Sinemurian and Pliensbachian stages of the Early Jurassic Period or approximately 199 to 182 million years ago.[4] This formation is part of the Glen Canyon Group that includes formations not only in northern Arizona but also parts of southeastern Utah, western Colorado, and northwestern New Mexico. The formation was primarily deposited by rivers. During the Early Jurassic period, the land that is now the Kayenta Formation experienced rainy summers and dry winters. By the Middle Jurassic period it was being encroached upon from the north by a sandy dune field that would become the Navajo Sandstone.[5] The animals here were adapted to a seasonal climate and abundant water could be found in streams, ponds and lakes.
Paleofauna
Oligokyphus lived beneath the feet of dinosaurs, such as the theropods Dilophosaurus, Kayentavenator[6] Megapnosaurus kayentakatae, the basal sauropodomorph Sarahsaurus,[7]heterodontosaurids, and the armored dinosaurs Scelidosaurus and Scutellosaurus. The Kayenta Formation has produced that remains of three coelophysoid taxa of different body size, which represents the most diverse ceratosaur fauna yet known.[8] The Kayenta Formation has yielded a small but growing assemblage of organisms. Vertebrates present here at the time of Oligokyphus included hybodont sharks, bony fish known as osteichthyes, lungfish, salamanders, the frog Prosalirus, the caecilian Eocaecilia, the turtle Kayentachelys, a sphenodontian reptile, various lizards. Also present were the synapsids Dinnebiton and Kayentatherium,[9] several early crocodylomorphs including Calsoyasuchus, Eopneumatosuchus, Kayentasuchus, and Protosuchus), and the pterosaur Rhamphinion.[9][10][11][12] The possible presence of the early true mammal Dinnetherium, and a haramyid mammal has also been proposed, based on fossil finds.[10] Vertebrate trace fossils from this area included coprolites[13] and the tracks of therapsids, lizard-like animals, and dinosaurs, which provided evidence that these animals were also present.[14] Non-vertebrates in this ecosystem included microbial or "algal" limestone,[13] freshwater bivalves, freshwater mussels and snails,[5] and ostracods.[15] The plant life known from this area included trees that became preserved as petrified wood.[11]
Reproduction
It is very likely that Oligokyphus had parental care. This is assumed to be true because most derived cynodonts have several traits associated with parental care in modern mammals.[16] Tritylodontids possess evidence of diphyodonty, a trait associated with suckling and therefore the production of milk.[17]
Oligokyphus, like other tritylodontids, possesses epipubic bones, which would have prevented the expansion of the torso and forced it to give birth to larval young like modern marsupials and monotremes.[18]
References
- Trevor Dykes. "JURASSIC CYNODONTS; Tritylodontidae, an internet directory". Archived from the original on 2010-04-23. Retrieved 8 June 2012.
- Sovak, J. 1999. Before the Dinosaurs (Coloring Book). Dover Publications. p.43 ISBN 0-486-40568-0
- J. M. Clark and D. E. Fastovsky. 1986. Vertebrate biostratigraphy of the Glen Canyon Group in northern Arizona. The Beginning of the Age of the Dinosaurs: Faunal change across the Triassic-Jurassic boundary, N. C. Fraser and H.-D. Sues (eds.), Cambridge University Press 285–301
- Padian, K (1997) Glen Canyon Group In: Encyclopedia of Dinosaurs, edited by Currie, P. J., and Padian, K., Academic Press.
- Harshbarger, J. W.; Repenning, C. A.; Irwin, J. H. (1957). Stratigraphy of the uppermost Triassic and the Jurassic rocks of the Navajo country. Professional Paper. 291. Washington, D.C.: U.S. Geological Survey.
- Gay, R. 2010. Kayentavenator elysiae, a new tetanuran from the early Jurassic of Arizona. Pages 27–43 in Gay, R. Notes on early Mesozoic theropods. Lulu Press (on-demand online press).
- Rowe, T. B.; Sues, H.-D.; Reisz, R. R. (2011). "Dispersal and diversity in the earliest North American sauropodomorph dinosaurs, with a description of a new taxon". Proceedings of the Royal Society B: Biological Sciences. 278 (1708): 1044–1053. doi:10.1098/rspb.2010.1867. PMC 3049036. PMID 20926438.
- Tykoski, R. S., 1998, The Osteology of Syntarsus kayentakatae and its Implications for Ceratosaurid Phylogeny: Theses, The University of Texas, December 1998.
- Jenkins, F. A. Jr.; Crompton, A. W.; Downs, W. R. (1983). "Mesozoic mammals from Arizona: new evidence in mammalian evolution". Science. 222 (4629): 1233–1235. doi:10.1126/science.222.4629.1233. PMID 17806725.
- Lucas, S. G.; Heckert, A. B.; Tanner, L. H. (2005). "Arizona's Jurassic fossil vertebrates and the age of the Glen Canyon Group". In Heckert, A. B.; Lucas, S. G. (eds.). Vertebrate paleontology in Arizona. Bulletin. 29. Albuquerque, NM: New Mexico Museum of Natural History and Science. pp. 95–104.
- Jenkins, F. A.; Jr; Shubin, N. H. (1998). "Prosalirus bitis and the anuran caudopelvic mechanism". Journal of Vertebrate Paleontology. 18 (3): 495–510. doi:10.1080/02724634.1998.10011077.
- Curtis, K.; Padian, K. (1999). "An Early Jurassic microvertebrate fauna from the Kayenta Formation of northeastern Arizona: microfaunal change across the Triassic-Jurassic boundary". PaleoBios. 19 (2): 19–37.
- Luttrell, P. R., and Morales, M. 1993. Bridging the gap across Moenkopi Wash: a lithostratigraphic correlation. Aspects of Mesozoic geology and paleontology of the Colorado Plateau. Pages 111–127 in Morales, M., editor. Museum of Northern Arizona, Flagstaff, AZ. Bulletin 59.
- Hamblin, A. H., and Foster, J. R. 2000. Ancient animal footprints and traces in the Grand Staircase-Escalante National Monument, south-central Utah. Pages 557–568 in Sprinkel, D. A., Chidsey, T. C., Jr., and Anderson, P. B. editors. Geology of Utah's parks and monuments. Utah Geological Association, Salt Lake City, UT. Publication 28.
- Lucas, S. G.; Tanner, L. H. (2007). "Tetrapod biostratigraphy and biochronology of the Triassic-Jurassic transition on the southern Colorado Plateau, USA". Palaeogeography, Palaeoclimatology, Palaeoecology. 244 (1–4): 242–256. doi:10.1016/j.palaeo.2006.06.030.
- Kielan-Jaworowska, Z; Luo, ZX; Cifelli, RL (2004). Mammals from the Age of Dinosaurs. Columbia University Press. Chapter 4. ISBN 978-0-231-11918-4.
- Hu, Yaoming; Meng, Jin; Clark, James M. (2009). "A New Tritylodontid from the Upper Jurassic of Xinjiang, China". Acta Palaeontologica Polonica. 54 (3): 385–391. doi:10.4202/app.2008.0053.
- Reily, Stephen; White, Thomas (January 2003). "Hypaxial Motor Patterns and the Function of Epipubic Bones in Primitive Mammals". Science. 299 (5605): 400–2. doi:10.1126/science.1074905. PMID 12532019.
- Hans-Dieter Sues. Dinnebitodon Amarall, a New Tritylodontid (Synapsida) from the lower Jurassic of Western North America. Journal of Paleontology. 60. pp. 758–762. JSTOR 1305061.
- Abdala, Fernando; Maria Claudia Malabarba (30 August 2007). "Enamel microstructure in Exaeretodon, a Late Triassic South American traversodontid (Therapsida: Cynodontia)". Revista Brasileira de Paleontologia. 10 (2): 71–78. doi:10.4072/rbp.2007.2.01.
- Johnson R. Haas. "the late Paleozoic land faunas" (PDF). GEOS 2000 Evolution of Life. Western Michigan University. Archived from the original (PDF) on 18 February 2012. Retrieved 8 June 2012.
External links
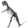 Paleontology portal
Paleontology portal- M. Alan Kazlev. "Tritylodontidae-Oligokyphus". Kheper – transformation – evolution – metamorphosis. Retrieved 8 June 2012.
- Michael R. Long. "Oligokyphus". Picture library. Natural History Museum (London). 001460. Archived from the original on 3 March 2016. Retrieved 8 June 2012.
- "Specimen account. Catalog Number: V83672". Berkeley Natural History Museums. Archived from the original on 12 December 2012. Retrieved 8 June 2012.








