Anchiornis
Anchiornis is a genus of small, four-winged paravian dinosaur. The genus Anchiornis contains only the type species Anchiornis huxleyi, named for its similarity to modern birds. Anchiornis fossils have been only found in the Tiaojishan Formation of Liaoning, China, in rocks dated to the Late Jurassic, about 160 million years ago. Anchiornis is known from hundreds of specimens, and given the exquisite preservation of some of these fossils, it became the first Mesozoic dinosaur species for which almost the entire life appearance could be determined, and an important source of information on the early evolution of birds.[1] Anchiornis huxleyi translates to "Huxley's near-bird" in Greek.
| Anchiornis | |
|---|---|
 | |
| Fossil specimen on display at the Beijing Museum of Natural History | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | Avialae (?) |
| Family: | †Anchiornithidae |
| Genus: | †Anchiornis Xu et al., 2009 |
| Species: | †A. huxleyi |
| Binomial name | |
| †Anchiornis huxleyi Xu et al., 2009 | |
Description

Anchiornis huxleyi was a small, bipedal theropod dinosaur with a triangular skull bearing several details in common with dromaeosaurids, troodontids, and primitive avialans. Like other early paravians, Anchiornis was small, about the size of a crow.[2] It had long, wing-bearing arms, long legs, and a long tail. Like all paravians, it was covered in feathers, though it also had scales on certain parts of the body. The wings, legs, and tail supported long but relatively narrow vaned feathers. Two types of simpler, downy (plumaceous) feathers covered the rest of the body, as in Sinornithosaurus: down feathers made up of filaments attached at their bases, and more complex down feathers with barbs attached along a central quill. Long, simple feathers covered almost the entire head and neck, torso, upper legs, and the first half of the tail. The rest of the tail bore pennaceous tail feathers (rectrices).[3] Long feathers on the head (crown) may have formed a crest.[1] While the first specimen of Anchiornis preserved only faint traces of feathers around the preserved portion of the body, many more well-preserved fossils have since been found. Studies of Anchiornis specimens using laser fluorescence have revealed not only more details of the feathers, but also of the skin and muscle tissue. Taken together, this evidence has given scientists a nearly complete picture of Anchiornis anatomy. Additional studies indicate that Anchiornis had body plumage that consisted of short quills with long and independent, flexible barbs. These barbs stuck out from the quills at low angles on two opposing blades. This also gave each feather an overall forked shape and resulted in the theropod possessing a softer textured and "shaggier" appearing plumage than is seen in modern birds. 'Shaggy' contour feathers probably influenced thermoregulatory and water repellence abilities, and, in combination with open-vaned wing feathers, would have decreased aerodynamic efficiency.[4][5]
Its size was originally estimated at 34 centimetres (13 in; 1.12 ft) and 0.11 kilograms (0.24 lb), but some specimens show that is could in fact have grown somewhat larger, up to 40 centimetres (16 in; 1.3 ft) and 0.25 kilograms (0.55 lb).[6] Anchiornis had a wing span of up to 50 centimetres (20 in; 1.6 ft).[7]
Wings

Like other early paravians, Anchiornis had large wings made up of pennaceous feathers attached to the arm and hand. The wing of Anchiornis was composed of 11 primary feathers and 10 secondary feathers. The primary feathers in Anchiornis were about as long as the secondaries, and formed a rounded wing. The wing feathers had curved but symmetrical central quills, with small and thin relative size, and rounded tips, all indicating poor aerodynamic ability. In the related dinosaurs Microraptor and Archaeopteryx, the longest wing feathers were closest to the tip of the wing, making the wings appear relatively long and pointed. However, in Anchiornis, the longest wing feathers were those nearest the wrist, making the wing broadest in the middle and tapering near the tip for a more rounded, less flight-adapted profile.[3]
Like other maniraptorans, Anchiornis had a propatagium, a flap of skin connecting the wrist to the shoulder and rounding out the front edge of the wing. In Anchiornis, this part of the wing was covered in covert feathers which smoothed the wing and covered the gaps between the larger primary and secondary feathers. However, unlike modern birds, the covert feathers of Anchiornis were not arranged in tracts or rows.[8] The arrangement of the covert feathers was also more primitive in Anchiornis than in birds and more advanced paravians. In modern birds, the coverts usually cover only the upper portion of the wing, with most of the wing surface made up of uncovered flight feathers. In some Anchiornis fossils, on the other hand, several layers of covert feathers seem to extend down to cover most of the wing's surface, so that the wing is essentially made of multiple layers of feathers, rather than a layer of broad feathers with only their bases hidden by layers of coverts. This multi-layered wing arrangement might have helped strengthen the wing, considering that the primary and secondary feathers themselves were narrow and weak.[9]
The wing included three clawed fingers; however, unlike in some more primitive theropods, the longest two fingers were not separate, but were bound together by the skin and other tissue forming the wing, so Anchiornis was functionally two-fingered.[8] These bound fingers were incorporated into a post-patagium, or flap of skin and other tissues that helped support the bases of the main wing feathers. Like the toes, the skin around the bottom of the fingers was covered in tiny, rounded scales. Unlike the toes, the flesh around the underside of the finger bones was twice as thick as the bones themselves and lacked distinct pads; instead, the fingers were straight and smooth without any major creases at the joints. Scales and skin around the fingers is very rarely preserved in fossils of early pennaraptorans, the only notable exceptions being Anchiornis and Caudipteryx, which had similar thick, scaly fingers associated with its wings.[8]
Legs
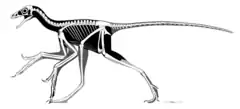
In addition to the front wings, Anchiornis had long, vaned feathers on the hind legs. This has led many scientists to call Anchiornis a four-winged dinosaur, along with similar animals like Microraptor and Sapeornis.[10] However, the feathers on the hind legs in Anchiornis did not have the shape or arrangement expected from flight feathers, and it is likely that their primary role was in display rather than flight.[11]
Anchiornis had very long legs, which is usually an indication that an animal is a strong runner. However, the extensive leg feathers indicate that this may be a vestigial trait, as running animals tend to have reduced, not increased, hair or feathers on their legs.[3] Like most paravians, Anchiornis had four toes on the foot, with the third and fourth toes the longest. The first toe, or hallux, was not reversed as in perching species.[3] The hindwings of Anchiornis were also shorter than those of Microraptor, and were made up of 12 to 13 flight feathers anchored to the tibia (lower leg) and 10 to 11 to the tarsus (upper foot). Also unlike Microraptor, the hindwing feathers were longest closer to the body, with the foot feathers being short and directed downward, almost perpendicular to the foot bones.[3]
Unlike many other paravians, the feet of Anchiornis (except for the claws) were completely covered in feathers, though these were much shorter than the ones making up the hindwing.[3] Some specimens have preserved scales on the toes, tarsus, and even lower leg (tibia), suggesting that scales existed beneath the feathers. The underside of the toes were formed into fleshy pads with distinct creases at the joints. The foot pads were covered in small, pebble-like scales. Scales were also present on the top of the feet but these are very hard to see in all known fossils.[8]
Color

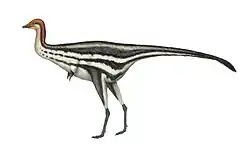
In 2010, a team of scientists examined numerous points among the feathers of an extremely well-preserved Anchiornis specimen in the Beijing Museum of Natural History to survey the distribution of melanosomes, the pigment cells that give feathers their color. By studying the types of melanosomes and comparing them with those of modern birds, the scientists were able to map the specific colors and patterning present on this Anchiornis when it was alive. Though this technique had been used and described for isolated bird feathers and portions of other dinosaurs (such as the tail of Sinosauropteryx), Anchiornis became the first Mesozoic dinosaur for which almost the entire life coloration was known (note that the tail of this specimen was not preserved).[1] The study found that most of the body feathers of this Anchiornis specimen were gray and black. The crown feathers were mainly rufous with a gray base and front, and the face had rufous speckles among predominantly black head feathers. The forewing and hindwing feathers were white with black tips. The coverts (shorter feathers covering the bases of the long wing feathers) were gray, contrasting the mainly white main wings. The larger coverts of the wing were also white with gray or black tips, forming rows of darker dots along mid-wing. These took the form of dark stripes or even rows of dots on the outer wing (primary feather coverts) but a more uneven array of speckles on the inner wing (secondary coverts). The shanks of the legs were gray other than the long hindwing feathers, and the feet and toes were black.[1]
In 2015, a second Anchiornis fossil at the Yizhou Fossil & Geology Park was subjected to a similar study that included a survey of melanosome shapes across all the feathers. In contrast to the 2010 study, only gray-black type melanosomes were found. Even when the crown feathers were examined, none of the rounder, rufous-type melanosomes were seen. The scientists who conducted this second study suggested several possible explanations for this discrepancy. First, the different preservation of melanosomes or different investigative techniques might have influenced the results of the original study. Second, because the Beijing Museum specimen was smaller, it is possible that the rufous color was replaced as these animals aged. Third, it is possible that there were regional differences or even different species of Anchiornis which had different color patterns in their plumage.[12]
Discovery and history

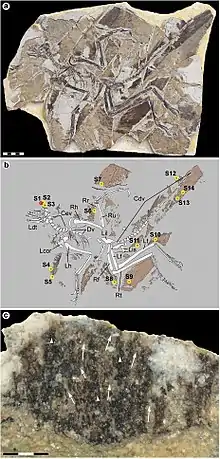
The first known fossil of Anchiornis (its type specimen) was dug up in the Yaolugou area of Jianchang County, Liaoning, China. These rocks have been difficult to date, but most studies have concluded that they belong to the Tiaojishan Formation of rocks dated to the late Jurassic period (Oxfordian age), 160.89 to 160.25 million years old.[3][13][14] studied and described by paleontologist Xu Xing and colleagues in a paper accepted to the Chinese Science Bulletin in 2009. The specimen is currently in the collection of the Institute of Vertebrate Paleontology and Paleoanthropology with the catalogue number IVPP V14378. It is an articulated skeleton missing the skull, part of the tail, and the right forelimb. The name Anchiornis huxleyi was chosen by Xu and colleagues in honor of Thomas Henry Huxley, an early proponent of biological evolution, and one of the first to propose a close evolutionary relationship between birds and dinosaurs. The generic name Anchiornis comes from combining the Ancient Greek words for "nearby" and "bird", because it was interpreted as important in filling a gap in the transition between the body plans of birds and dinosaurs.[15]
A second specimen came to light around the same time, and was given to a team of scientists from Shenyang Normal University by a local farmer when they were excavating at the Yaolugou dig site. According to the farmer, this second specimen had been found nearby in the area of Daxishan, also from Tiaojishan Formation rocks of about the same age as the first Anchiornis. Two scientists visited the site in order to compare the new fossil with the rock types found there, and were able to confirm that the new specimen probably did come from the area the farmer described. They were able to dig up several fish fossils and a third Anchiornis fossil. The farmer's fossil underwent study which was published on September 24, 2009, in the journal Nature. It was assigned the catalogue number LPM – B00 169 in the Liaoning Paleontological Museum. It is larger and much more complete than the first specimen, and preserved long wing feathers on the hands, arms, legs and feet, showing that it was a four-winged dinosaur similar to Microraptor.[8]

While only a few specimens have been described in detail, many more have been identified and are held in both private collections and museums. One of these, a nearly complete skeleton missing the tail, also preserving extensive feather remains, was reported in 2010. This fossil also showed evidence that Anchiornis had a feathered crest on its head, and was used to determine the animal's life coloration. It is housed in the Beijing Museum of Natural History with the specimen number BMNHC PH828.[1] Another specimen from the same fossil quarry as the type specimen was found by a local fossil dealer and sold to the Yizhou Fossil & Geology Park and catalogued there as YTGP-T5199. This fossil, a nearly complete skeleton, was prepared and studied by scientists at the Geology Park and identified as an Anchiornis. It was then used for a scanning electron microscope study of Anchiornis feather microstructure. The study also examined the well-preserved melanosomes of the feathers to determine their color. The scientists involved in the study found that the coloration found for this specimen was different than the color reported for BMNHC PH828, and they noted that the BMNHC specimen may not in fact be Anchiornis, as it was described before similar species from the same formation had been discovered.[12]
The Shandong Tianyu Museum of Nature in Pingyi County, China, for example, was reported to hold 255 specimens of Anchiornis in its collections in 2010.[16] Among their collection is a very well preserved fossil with visible color patterns, catalogued as STM 0-214. While this specimen has yet to be fully described, it was photographed for a 2011 article in National Geographic and was used in a study of Anchiornis covert feathers and wing anatomy the following year.[9]
Classification
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Cladogram following the results of a phylogenetic study by Lefèvreet al., 2017.[17] |
When it was first discovered, the scientists who studied Anchiornis conducted a phylogenetic analysis and concluded that it was an early member of the group Avialae, along with Archaeopteryx.[15] Members of Avialae, called avialans, are all more closely related to modern birds than they are to dromaeosaurid and troodontid dinosaurs, though the earliest and most primitive members of all three groups are extremely similar to each other, which makes it difficult to sort out exactly which of these three main paravian branches they belong to.[2]
The second specimen of Anchiornis was more complete than the first, and preserved several features which led Hu Dongyu and his colleagues to reclassify Anchiornis as a troodontid.[3] Several more studies using similar analyses have also found Anchiornis to be a troodontid, though there have been exceptions. One study found Anchiornis to be a member of Archaeopterygidae, and it along with Archaeopteryx were considered more primitive than dromeosaurids, troodontids, or avialans. In 2015, Sankar Chatterjee placed Anchiornis along with Microraptor and other four-winged paravians in a group he called "Tetrapterygidae", just outside the Avialae, though this was not supported with a phylogenetic analysis.[18] More comprehensive studies suggested that Anchiornis may have been an avialan after all,[19] though new finds and updated versions of the same study later reversed this finding, concluding that Anchiornis was most likely a basal member of the clade Paraves, just outside the clade that includes dromaeosaurids, troodontids, and avialans.[17]
In a 2017 re-evaluation of the Haarlem Archaeopteryx specimen, Anchiornis was found to be in a group with other genera, like Eosinopteryx, Xiaotingia, and was placed in the family Anchiornithidae along with other relatives.[20][21]
Paleobiology
Anchiornis is notable for its proportionally long forelimbs, which measured 80% of the total length of the hindlimbs. This is similar to the condition in early avians such as Archaeopteryx, and the authors pointed out that long forelimbs are necessary for flight. Anchiornis also had a more avian wrist than other non-avialan theropods. The authors initially speculated that it would have been possible for Anchiornis to fly or glide. However, further finds showed that the wings of Anchiornis, while well-developed, were short when compared to later species like Microraptor, with relatively short primary feathers that had rounded, symmetrical tips, unlike the pointed, aerodynamically proportioned feathers of Microraptor.[3] A 2016 study of potential flight performance in early paravians concluded that while juvenile Anchiornis specimens may have been able to use their wings to assist running up an incline, and could possibly have achieved flapping flight if a very high-angle flapping wing stroke was used, the larger adult specimens would not have gained any aerodynamic benefit from their wings—they were simply too heavy compared to their total wing area. The same study found that flapping the wings while running would have resulted in a small (10%) increase to its running speed. Similarly, use of the wings during leaping would have resulted in a 15 to 20% increase in height and distance.[7] Notably, Anchiornis seems to have lacked a breastbone (sternum), which may have been made of cartilage rather than bone, as in more primitive theropods.[22][23]
.jpg.webp)
Anchiornis has hindleg proportions more like those of more primitive theropod dinosaurs than avialans, with long legs indicating a fast-running lifestyle. However, while long legs normally indicate a fast runner, the legs and even feet and toes of Anchiornis were covered in feathers, including long feathers on the legs, similar to those in the hindwings of Microraptor.[3] Long leg feathers on the lower legs may have slowed the running speed of Anchiornis. In modern birds, especially those that live on the ground, the lower legs tend to show reduction or even loss of feathers.[3] The hind wings of Anchiornis were smaller and made of more curved, symmetrical feathers than those of Microraptor, suggesting that they were used mainly for display rather than flight.[11] However, they might still have granted the animal some kind of aerodynamic advantage, even if their primary purpose was for display or some other function.[2][24]
The skeletal structure of Anchiornis is similar to Eosinopteryx, which appears to have been a capable runner due to its uncurved rear claws and absence of flight feathers on its tail or lower legs. Anchiornis shared a similar body plan and the same ecosystem as Eosinopteryx, suggesting different niches and a complex picture for the origin of flight.[25][26]
Like many modern birds, Anchiornis exhibited a complex pattern of coloration with different colors in speckled patterns across the body and wings, or "within- and among-feather plumage coloration." In modern birds, such color patterning is used in communication and display, either to members of the same species (e.g. for mating or territorial threat display) or to threaten and warn off competing or predatory species.[1]
Feeding
A 2018 study reported gastric pellets in association with Anchiornis specimens; some of the Anchiornis were even preserved with pellets still inside their bodies. Anchiornis is the earliest theropod known to have produced pellets. The pellets contained lizard bones and ptycholepid fish scales.[27]
See also
References
- Li, Q.; Gao, K.-Q.; Vinther, J.; Shawkey, M. D.; Clarke, J. A.; d'Alba, L.; Meng, Q.; Briggs, D. E. G.; Prum, R. O. (2010). "Plumage color patterns of an extinct dinosaur" (PDF). Science. 327 (5971): 1369–1372. Bibcode:2010Sci...327.1369L. doi:10.1126/science.1186290. PMID 20133521. S2CID 206525132.
- Witmer, L. M. (2009). "Palaeontology: Feathered dinosaurs in a tangle". Nature. 461 (7264): 601–602. Bibcode:2009Natur.461..601W. doi:10.1038/461601a. PMID 19794481. S2CID 205049989.
- Hu, D.; Hou, L.; Zhang, L.; Xu, X. (2009). "A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus". Nature. 461 (7264): 640–643. Bibcode:2009Natur.461..640H. doi:10.1038/nature08322. PMID 19794491. S2CID 205218015.
- "Feathered dinosaurs were even fluffier than we thought". sciencedaily.com.
- Saitta, Evan T.; Gelernter, Rebecca; Vinther, Jakob (2018). "Additional information on the primitive contour and wing feathering of paravian dinosaurs". Palaeontology. 61 (2): 273–288. doi:10.1111/pala.12342. hdl:1983/61351c6d-1517-4101-bac8-50cbb733761d.
- Paul, G.S. (2010). The Princeton Field Guide to Dinosaurs. Princeton University Press. p. 128.
- Dececchi, T.A.; Larsson, H.C.E.; Habib, M.B. (2016). "The wings before the bird: an evaluation of flapping-based locomotory hypotheses in bird antecedents". PeerJ. 4: e2159. doi:10.7717/peerj.2159. PMC 4941780. PMID 27441115.
- Wang, X.; Pittman, M.; Zheng, X.; Kaye, T.G.; Falk, A.R.; Hartman, S.A.; Xu, X. (2017). "Basal paravian functional anatomy illuminated by high-detail body outline". Nature Communications. 8: 14576. Bibcode:2017NatCo...814576W. doi:10.1038/ncomms14576. PMC 5339877. PMID 28248287.
- Longrich, N.R.; Vinther, J.; Meng, Q.; Li, Q.; Russell, A.P. (2012). "Primitive wing feather arrangement in Archaeopteryx lithographica and Anchiornis huxleyi". Current Biology. 22 (23): 2262–2267. doi:10.1016/j.cub.2012.09.052. PMID 23177480.
- Xu, X.; Zhou, Z.; Dudley, R.; Mackem, S.; Chuong, C.M.; Erickson, G.M.; Varricchio, D.J. (2014). "An integrative approach to understanding bird origins". Science. 346 (6215): 1253293. doi:10.1126/science.1253293. PMID 25504729. S2CID 24228777.
- O'Connor, J.K.; Chang, H. (2015). "Hindlimb feathers in paravians: primarily "wings" or ornaments?". Biology Bulletin. 42 (7): 616–621. doi:10.1134/s1062359015070079. S2CID 15637177.
- Lindgren, J.; Sjövall, P.; Carney, R. M.; Cincotta, A.; Uvdal, P.; Hutcheson, S. W.; Gustafsson, O.; Ulysse Lefèvre, U.; François Escuillié, F.; Heimdal, J.; Anders Engdahl, A.; Gren, J.A.; Kear, B.P.; Wakamatsu, K.; Yans, J.; Godefroit, P. (2015). "Molecular composition and ultrastructure of Jurassic paravian feathers". Scientific Reports. 5: 13520. doi:10.1038/srep13520. PMC 4550916. PMID 26311035.
- Liu, Y.-Q.; Kuang, H.-W.; Jiang, X.-J.; Peng, N.; Xu, H.; Sun, H.-Y. (2012). "Timing of the earliest known feathered dinosaurs and transitional pterosaurs older than the Jehol Biota". Palaeogeography, Palaeoclimatology, Palaeoecology. 323–325: 1–12. doi:10.1016/j.palaeo.2012.01.017.
- Chu, Z.; He, H.; Ramezani, J.; Bowring, S.A.; Hu, D.; Zhang, L.; Zheng, S.; Wang, X.; Zhou, Z.; Deng, C.; Guo, J. (2016). "High-precision U-Pb geochronology of the Jurassic Yanliao Biota from Jianchang (western Liaoning Province, China): Age constraints on the rise of feathered dinosaurs and eutherian mammals". Geochemistry, Geophysics, Geosystems. 17 (10): 3983. Bibcode:2016GGG....17.3983C. doi:10.1002/2016GC006529.
- Xu, X; Zhao, Q.; Norell, M.; Sullivan, C.; Hone, D.; Erickson, G.; Wang, X.; Han, F.; Guo, Y. (2009). "A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin". Chinese Science Bulletin. 54 (3): 430–435. doi:10.1007/s11434-009-0009-6.
- "China has world's largest dinosaur museum". Dawn. 4 August 2010. Archived from the original on 2010-10-28. Retrieved 2010-11-06.
- Lefèvre, Ulysse; Cau, Andrea; Cincotta, Aude; Hu, Dongyu; Chinsamy, Anusuya; Escuillié, François; Godefroit, Pascal (2017). "A new Jurassic theropod from China documents a transitional step in the macrostructure of feathers". The Science of Nature. 104 (9–10): 74. Bibcode:2017SciNa.104...74L. doi:10.1007/s00114-017-1496-y. PMID 28831510. S2CID 32780661.
- Chatterjee, S. (2015). The Rise of Birds: 225 million Years of Evolution. (2nd edition). Johns Hopkins University Press.
- Godefroit, P.; Cau, A.; Dong-Yu, H.; Escuillié, F.; Wenhao, W.; Dyke, G. (2013). "A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds". Nature. 498 (7454): 359–362. Bibcode:2013Natur.498..359G. doi:10.1038/nature12168. PMID 23719374. S2CID 4364892.
- Foth, C.; Rauhut, O.W.M. (2017). "Re-evaluation of the Haarlem Archaeopteryx and the radiation of maniraptoran theropod dinosaurs". BMC Evolutionary Biology. 17 (1): 236. doi:10.1186/s12862-017-1076-y. PMC 5712154. PMID 29197327.
- Cau, A.; Beyrand, V.; Voeten, D.; Fernandez, V.; Tafforeau, P.; Stein, K.; Barsbold, R.; Tsogtbaatar, K.; Currie, P.; Godefroit, P. (2017). "Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs". Nature. 552 (7685): 395–399. Bibcode:2017Natur.552..395C. doi:10.1038/nature24679. PMID 29211712. S2CID 4471941.
- Zheng, X.; O'Connor, J.; Wang, X.; Wang, M.; Zhang, X.; Zhou, Z. (2014). "On the absence of sternal elements in Anchiornis (Paraves) and Sapeornis (Aves) and the complex early evolution of the avian sternum". Proceedings of the National Academy of Sciences. 111 (38): 13900–13905. doi:10.1073/pnas.1411070111. PMID 25201982.
- Foth, C (2014). "Comment on the absence of ossified sternal elements in basal paravian dinosaurs". Proceedings of the National Academy of Sciences. 111 (50): E5334. doi:10.1073/pnas.1419023111. PMC 4273336. PMID 25480553.
- Zheng, X.; Zhou, Z.; Wang, X.; Zhang, F.; Zhang, X.; Wang, Y.; Wei, G.; Wang, S.; Xu, X. (2013). "Hind wings in basal birds and the evolution of leg feathers". Science. 339 (6125): 1309–1312. Bibcode:2013Sci...339.1309Z. doi:10.1126/science.1228753. PMID 23493711. S2CID 206544531.
- Godefroit, P.; Demuynck, H.; Dyke, G.; Hu, D.; Escuillié, F.; Claeys, P. (2012). "Reduced plumage and flight ability of a new Jurassic paravian theropod from China". Nature Communications. 4: 1394. Bibcode:2013NatCo...4.1394G. doi:10.1038/ncomms2389. PMID 23340434.
- Fraser, G. (2014). ""Bizarre structures" point to dromaeosaurs as parasites and a new theory for the origin of avian flight" (Automatic PDF download). Journal of Paleontological Sciences. 6: 1–27.
- Xu, X.; Li, F.; Wang, Y.; Sullivan, C.; Zhang, F.; Zhang, X.; Sullivan, C.; Wang, X.; Zheng, X. (2018). "Exceptional dinosaur fossils reveal early origin of avian-style digestion". Scientific Reports. 8 (1): 14217. Bibcode:2018NatSR...814217Z. doi:10.1038/s41598-018-32202-x. ISSN 2045-2322. PMC 6155034. PMID 30242170.




.png.webp)



.jpg.webp)





