Achillobator
Achillobator (/əˌkɪləˈbeɪtɔːr/ ə-KIL-ə-BAY-tor; meaning "Achilles hero") is a genus of large dromaeosaurid dinosaur that lived in Asia during the Late Cretaceous period about 96 million to 89 million years ago in what is now the Bayan Shireh Formation. The genus is currently monotypic, only including the type species A. giganticus. The first remains were found in 1989 during a Mongolian-Russian field expedition in Mongolia and later described in 1999. Since the first discovery and description, no more specimens have been found. It represents the first and largest dromaeosaurid known from the Bayan Shireh Formation.
| Achillobator | |
|---|---|
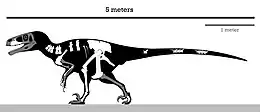 | |
| Skeletal diagram showing the size and preserved elements from holotype | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Family: | †Dromaeosauridae |
| Clade: | †Eudromaeosauria |
| Subfamily: | †Dromaeosaurinae |
| Genus: | †Achillobator Perle et al. 1999 |
| Type species | |
| †Achillobator giganticus Perle et al. 1999 | |
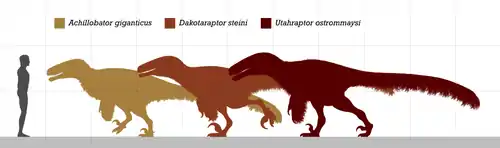
It was a large, heavy-built, ground-dwelling, bipedal carnivore that would have been an active feathered predator hunting with the enlarged sickle claw on each second toe. Estimated at 5 m (16 ft) in length with a weight about 250–348 kg (551–767 lb), Achillobator is considered to be one of the largest dromaeosaurs, along with Austroraptor, Dakotaraptor and Utahraptor. Achillobator was a deep-bodied and relatively short-armed dromaeosaurid with stocky and robust hindlimbs. Some of the most notable features consisted in the robustly built skeleton—an unusual trait in dromaeosaur dinosaurs, which were generally lightly built animals—such as the deep maxilla and femur, and the primitive pelvis, having a vertically oriented pubis that differs from the rest of dromaeosaurids.
Achillobator is classified as a dromaeosaurid taxon, more specifically within the Eudromaeosauria, a group of hypercarnivore dromaeosaurids that were mainly terrestrial instead of arboreal or amphibious. In most cladistic analyses Achillobator is recovered as a close relative of Dromaeosaurus and Utahraptor, although it is often considered to be the sister taxon of the latter. The stocky and short hindlimb ratio of Achillobator indicates that it was not cursorial—an animal adapted for speed or to maintain high speeds—moreover, the robust morphology of the maxilla suggests a predatory behavior based on large-sized prey.
History of discovery
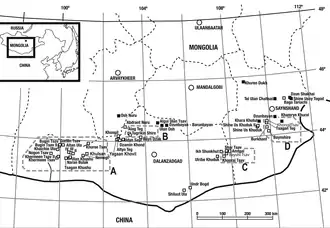
In 1989 during a field exploration examining the outcrops at the Khongil locality in South Central Mongolia, conducted by the Mongolian and Russian Paleontological Expedition in the Gobi Desert, many dinosaur fossil discoveries were made. About 5.6 km (5,600 m) away from this locality, a large and associated, but mostly disarticulated partial theropod skeleton was discovered in sediments of the Burkhant locality, Bayan Shireh Formation; no other findings were made by the expedition at this locality. It was found in fine-grained, medium sandstone/gray mudstone that was deposited dating back to the Late Cretaceous epoch. The specimen was found preserving a left maxilla with nine teeth and two empty alveoli, four cervical vertebrae, three dorsal vertebrae and eight caudal vertebrae, a nearly complete pelvic girdle compromising both pubes, right illium and right ischium, both femora and left tibia, left metatarsals III and IV, manual and pedal phalanges with some unguals, right scapulocoracoid, an isolated radius, two ribs and caudal chevrons. It was collected and prepared by the assistant paleontologist Namsarai Batulseen and stored as MNUFR-15. Ten years later, the specimen was formally described in 1999 and became the holotype for the new genus and species Achillobator giganticus. It was identified as a dromaeosaurid taxon. The description was performed by the Mongolian paleontologist Altangerel Perle, and North American paleontologists Mark A. Norell and James M. Clark. In terms of etymology, the generic name, Achillobator, is derived from the Latin word "Achillis" (genitive singular of Achilles) in reference to the large Achilles tendon that supported the second pedal ungual (known as "sickle claw") of most dromaeosaurids, and the old Mongolian word "баатар" (baatar, meaning hero).[1]
However, the description was published in a very preliminary format, being not complete at all, having issues with preserved elements and numerous typographical errors.[1] Due to a misinterpretation, the pedal ungual II (or sickle claw) was claimed to be preserved and to articulate with the pedal phalanx II, however, this was corrected by Senter in 2007 and this ungual actually represents a manual one.[2] Turner and colleagues in 2012 during their large revision of the Dromaeosauridae stated that the describing paper of Achillobator was likely published without the knowledge of the two latter paleontologists as indicated by a draft left in Mongolia in 1997.[3]
Chimera hypothesis
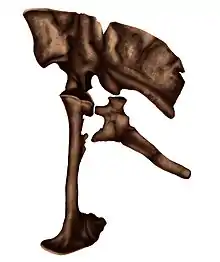
The pelvic girdle of Achillobator features plesiomorphic (primitive) saurischian characteristics compared to other dromaeosaurids. For instance, the pubis is aligned vertically and has a relatively large pubic boot (a wide expansion at the end), unlike most other dromaeosaurids, where there is generally a much smaller boot. The preserved vertebrae are very robust and features a series of pleurocoels. The above differences led Burnham and team in 2000 to suggest that the holotype of Achillobator in fact, represents a paleontological chimera, and only the pedal unguals may have come from a dromaeosaurid-grade dinosaur.[4]
However, given that the specimen was actually found in semiarticulation,[5] and all elements have the same color and preservation quality the assignment of remains to a single individual is supported. Despite the fact that Achillobator features unusual and primitive characteristics compared to other dromaeosaurids, it is commonly recovered as a taxon falling within Dromaeosauridae in cladistic analyses.[3]
Description

The holotype is estimated to have reached a length of 5 m (16 ft) and weighed 250–348 kg (551–767 lb).[6][7] Achillobator represents the second largest described dromaeosaurid taxon (after Utahraptor) with a tibial length of 49 cm (490 mm).[3] The gigantic size of Achillobator was only approached or surpassed by the also paravians Austroraptor, Dakotaraptor and Utahraptor.[8] The holotype was found lacking traces of feather integument,[1] however, there are strong evidence coming from other relatives that suggest the likely presence of plumage on Achillobator.[9][10]
According to the revised diagnosis by Turner and colleagues in 2012, Achillobator can be distinguished based on the following combination of characteristics and autapomorphies: the promaxillary fenestra is completely exposed; the promaxillary and maxillary fenestrae are elongate and vertically oriented at same level in the maxilla; metatarsal III is wide on the upper end; the femur is longer than the tibia; the pelvis is propubic (pubis extends forward towards the head of the animal); the obturator process on the ischium is large and triangular situated on the upper half of ischial shaft; and the boot at distal symphysis of pubis is developed in a cranial and caudal aspect.[3]
Skull
The maxilla was deep and robust, and measured about 29 cm (290 mm) in length. Its lateral side was smooth compared to the dorsal areas, but had a robust constitution. It had approximately 11 alveoli (sockets that held the roots of teeth). The teeth of Achillobator displayed marked homodonty (teeth of similar shape and size) and they were serrated and recurved with the posterior serrations being slightly larger than the anterior serrations. Along with the posterior serrations, there were 15 or 18 denticles per 5 mm (0.50 cm) near the center of the teeth, although the anterior serrations had 17 to 20 denticles per 5 mm (0.50 cm). The dimensions of the maxilla suggest that Achillobator had a relatively large skull competing to those of carnosaurs.[1]
Postcranial skeleton

The vertebral column was composed of large and robust cervical, dorsal and caudal vertebrae. The anterior series of cervical vertebrae were sharply-angled developed, with the anterior articulation facet of the centrum developed over the posterior facet. These characteristics are interpreted as indicators of a S-shaped neck, as seen in most maniraptorans. The dorsal vertebrae were stocky and had pleurocoels at the lateral surfaces, increasing in posterior vertebrae. Their central articulation facets were rounded and concave in shape. The neural processes of the dorsal vertebrae show resemblance to those of large ratites such as the extinct moa and extant emu, ostrich and cassowary, and they had protruding interspinous ligaments scars that indicates a robust and similar back to those birds. In all caudal vertebrae the centrum was amphiplatian (flat on both ends) with the posterior articulation facet being more concave than the anterior one. As indicated by a transitional caudal, the anterior neural spines were elongated and progressively disappeared on posterior caudals. The posterior series of caudals were articulated with long chevrons and had very elongated prezygapophyses (projections of the vertebral arch that connect adjacent vertebrae).[1]
The scapulocoracoid was formed by the fusion of the scapula and coracoid. In the posterior area of the acromion process, the scapula had a small tubercle that attached the area for the muscle scapulohumeralis in life. The glenoid was located at the bottom area facing to the lateral side. The scapula was relatively elongated and flattened at the scapular blade. The blade is thickened and slightly curved to the inner side following the rounded shape of the rib cage. The coracoid is robust and broad, measuring 16.7 cm (167 mm) long and 14.8 cm (148 mm) in height. The anterior edge had a coarse texture, likely for the articulation with cartilaginous tissue. A large foramen was located from the front to the back, probably acting as a channel for blood vessels and nerves. The radius measured 26 cm (260 mm) long.[1]
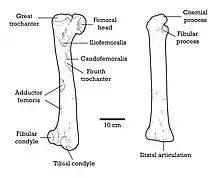
The femur was highly robust and longer than the tibia, a rare trait in dromaeosaurids, measuring 50.5 cm (505 mm) in length. On the anterior inner surface, there was a large, sculptured and concave surface that likely worked as the insertion for the m. iliofemoralis internus in life, and the lesser trochanter had the insertion for the m. pubo ischio femoralis on the lower edge. On the upper lateral surface of the femur, bottom-oriented to the greater trochanter, there was a tubercle that formed the attachment for the m. iliofemoralis. Insertions for m. ilio trochantericus and ischio trochantericus were located on the posterolateral surface at the proximal end of the femur. In the inner side, on the proximal third part of the shaft, a coarse, concave area likely attached m. pubo ischio femoralis externus-3. Below this area, a small, convex tubercle attended the insertion for m. pubo ischio femoralis-2. The tibia was less robust than the femur but more straight, measuring 49 cm (490 mm) in length. The surface of the shaft center was smooth, without traces of muscles. In front and towards the back surface of the cnemial crest however, there was a coarse and microsculptured area that worked as the attachment for m. ilio tibialis and ambiens. Additional tissue traces in the tibia were located on the lower surface and towards the posterior aspect. Here, a depression with a somewhat coarse surface likely attached prominent aponeurosis of the connected ligaments to the tibia and fibular head in life.[1]
Metatarsals III and IV were stocky, measuring 23.4 cm (234 mm) and 20.9 cm (209 mm) respectively, indicating that the length of the metatarsus was under the 50% of the tibial length. As in other maniraptorans, the metatarsals were closely united. The top end of metatarsal III was side to side flattened and not pinched at the upper end, thus lacking an arctometatarsalian (pinched upper end) condition. The lower end of metatarsal IV had a joint where motion was restricted to one plane, and the lateral condyle was short. The pelvis was formed by the pubis, ilium and ischium bones. The ilium was prominently tall measuring 51.3 cm (513 mm) in length and 28.8 cm (288 mm) in height with the preacetabular process situated from top to bottom. Both insertions for the m. caudofemoralis were located at the inner surface of the ilium. On the anterior edge were microstriations that likely gave form to the m. iliofemoralis in life, behind to this area, traces of the m. ilio tibialis-2 were also present. The pubis measured 54.8 cm (548 mm) long and is very straight having a large pubic boot (a wide expansion at the end). While other dromaeosaurids had a very opisthopubic pelvic configuration, Achillobator had a primitive propubic configuration. The large pubic boot had coarse areas on the dorsal surface that served as attachment sites for the m. pubo ischio femoralis internus ventralis. Its lateral surface was very flat and had numerous microstriations that probably originated the m. pubo ischio femoralis externus. The ischium was shorter than the ilium and pubis, measuring 37.8 cm (378 mm) in length. A large ridge was formed on its shaft and expanded to the anterior edge, likely forming the m. adductor femoris in life. This ridge-like structure was heavy-built compared to other dromaeosaurids. At the lateral sides the surface was very rough, possibly attaching m. flexor tibialis internus-1 in life.[1]
Classification

In its original description, Achillobator was placed as a close relative of Dromaeosaurus with an ambiguous position in the family.[1] In more recent and solid research Achillobator is classified within the Dromaeosauridae, a group of very bird-like, maniraptoran dinosaurs, being placed in the Eudromaeosauria, a group of dromaeosaurids that were obligate terrestrial and hypercarnivore animals, better known as the "true dromaeosaurids".[11] They strongly differ from other dromaeosaurs, such as the arboreal microraptorians[12] or amphibious halszkaraptorines.[13]
Eudromaeosauria was first defined as a node-based clade by Nicholas R. Longrich and Philip J. Currie in 2009 as the most inclusive natural group containing Dromaeosaurus, Velociraptor, Deinonychus, and Saurornitholestes, their most recent common ancestor and all of its other descendants. The various "subfamilies" have also been re-defined as clades, usually defined as all species closer to the groups namesake than to Dromaeosaurus or any namesakes of other sub-clades.[11]
The recent phylogenetic analysis conducted by Hartman and colleagues in 2019, recovers Achillobator as the sister taxon of Yixianosaurus.[14] Currie and Evans 2020 have recover the position of the genus within the Velociraptorinae.[15] Although some analyses have place the genus in unusual positions, Achillobator has been recovered as a dromaeosaurine very related to Utahraptor and Dromaeosaurus in most phylogenetic analyses.[16][3][17][13] Even more similarities within Achillobator and Utahraptor were found with the osteological reexamiation of Utahraptor specimens, supporting their close relationships.[18]
During the description of Halszkaraptor in 2017 and the coined Halszkaraptorinae, new affinities for dromaeosaurids were proposed in order to include this new subfamily. Achillobator was recovered in a basal position outside a clade formed by Dromaeosaurus and Utahraptor.[13] But in the results for the Eudromaeosauria phylogeny based on the phylogenetic analysis conducted by Jasinski and colleagues in 2020 during the description of Dineobellator, Achillobator was again recovered in the traditional Utahraptor+Achillobator clade but this time outside the Dromaeosaurinae.[19]
Left cladogram as in Cau et al. 2017,[13] and right cladogram in Jasinski et al. 2020:[19]
|
|
Paleobiology
Perle and team pointed out in 1999 that the structure of the hindlimbs and pelvic region of Achillobator indicates that the animal had massive thighs and robustly built legs suited for moderate fast-running. In addition, the prominent pelvis of Achillobator has its own specialized femoral muscle retractors that may indicate a strong ability for leaping.[1]
In 2016, Scott Persons IV and Currie examined the limb proportion of numerous theropods and found that compsognathids, troodontids and tyrannosauroids were cursorial animals with many taxa recovered with relatively high CLP (cursorial-limb-proportion) scores. Dromaeosaurids however, were recovered with low CLP scores, with Achillobator scoring −5.3 suggesting that it was not adapted to maintain high speeds for extended amounts of time.[20]
Predatory behavior
.jpg.webp)
In 2009 Longrich and Currie suggested that while other dromaeosaurids filled a variety of specialized ecological niches like the slender unenlagiines, eudromaeosaurs, such as Achillobator, retained a conservative life-style and filled the niche of large-bodied predators of often medium to large-sized prey.[11]
Manning and team in 2009 tested the function of the sickle claw of dromaeosaurids by analyzing the biomechanics of how stresses and strains would be distributed along the claws and into the limbs, and using comparisons within the curvature of the dromaeosaurid sickle claw on the foot with curvature in modern birds and mammals. They found that they were ideal for climbing and for a ground-dwelling life style.[21] Peter Mackovicky stated that the analysis might be correct on primitive dromaeosaurids (such as Microraptor) being tree-climbers, however, this does not explain why giant animals like Achillobator or Utahraptor retained sickle claws when they were way too large to have climbed trees. Mackovicky suggested that larger dromaeosaurids adapted the claw to be used exclusively for a more aggressive predatory behavior.[22]
The striking resemblance between the feet and legs of dromaeosaurids and those of accipitrid birds of prey, led Fowler et al. 2011 to propose that dromaeosaurids hunted in a similar way to those raptorial birds. They found that the feet and legs of dromaeosaurs resemble those of eagles and hawks by having an enlarged second claw and a similar range of flexion, but the metatarsals share more resemblance to those of owls. The model RPR (Raptor Prey Restraint), proposes that dromaeosaurs leaped into their prey, immobilizing it with their body weight, and then hold it tightly with the large, sickle-shaped claws; afterwards, the dromaeosaur would start to feed on the animal while it's still alive and the death will eventually came from blood loss and organ failure. The arms or "wings", that were likely covered in long feathers, may have been flapped by the dromaeosaur in order to stabilize its balance while restraining prey, along with this, the long, feathered tail probably worked as a counter-balance to the main body. Lastly, the snout would have been useful for finishing off its prey. With these observations, they established that dromaeosaurids and troodontids were niche partitioned as large and small prey predators, respectively.[23]
In 2020 Powers and colleagues re-examined the maxillae of eudromaeosaur taxa concluding that Asian and North American eudromaeosaurs were separated by snout morphology and ecological strategies. They found the maxilla to be a reliable reference when inferring the shape of the premaxilla and overall snout. For instance, most Asian species have elongated snouts based on the maxilla (animals like Velociraptor are known from complete skulls), indicating a selective feeding, such as picking up small, fast prey. Achillobator however, is an exception for Asian eudromaeosaurs, featuring a robust and deep maxillar morphology similar to North American eudromaeosaurs which also have stocky and deep snouts, and thus indicating relationships with these members. The adaptations of Achillobator and North American eudromaeosaurs indicate a diet based on large-sized prey.[24]
Paleoenvironment

The remains of Achillobator were unearthed from the Burkhant locality of the Bayan Shireh Formation. This formation has been divided into upper and lower parts, with some localities representing Lower and Upper Cretaceous boundary.[25][26][27][28] Burkhant, the type locality of Achillobator, has been identified as a Upper Cretaceous-boundary locality.[29] Analyses on the magnetostratigraphy of the formation indicate that the entire Bayan Shireh lies within the Cretaceous Long Normal, which lasted only until the end of the Santonian stage, giving a Cenomanian-Santonian age.[30] The recent calcite U–Pb analyses performed by Kurudama and colleagues in 2020 have confirm the age of the Bayan Shireh Formation from 95.9 ± 6.0 million to 89.6 ± 4.0 million years ago, also supporting a Cenomanian-Santonian age.[31] Over the time, a strong correlation with the Iren Dabasu Formation has been proposed by numerous authors mainly based on the similar fossil assemblages.[32][33][28] This hypothesis may be also supported by the similar ages.[34]
The environments that were present in the Bayan Shireh Formation and Achillobator inhabited were relatively humid and had extensive fluvial and lacustrine facies, that is, a well-watered region dominated by prominent meanders, rivers, lakes and streams. The climate of the formation was slightly semi-arid, as seen on caliche-based sediments.[32][30] Angiosperms were largely present in the formation, based on fossils of cornaceans and fossilized fruits at several localities.[25][35]
Contemporary paleofauna

Achillobator shared its surroundings in the Bayan Shireh Formation with other paleofauna such as the medium-sized theropods Erlikosaurus and Segnosaurus,[36] Garudimimus,[37] and possibly Alectrosaurus;[38] the heavy-built ankylosaurs Talarurus and Tsagantegia;[39] the small marginocephalians Amtocephale[40] and Graciliceratops;[41] the hadrosauroid Gobihadros;[29] and the large, long-necked sauropod Erketu.[35] Non-dinosaur taxa was present as well, mostly compromising semiaquatic and terrestrial reptiles like the turtles Lindholmemys, Gobiapalone and "Trionyx";[42][43] and the crocodylomorph Paralligator.[44] Fishes are also known from the formation, such as the shark Hybodus and the remains of osteichthyans at various localities.[28] Mammals were extremely rare around the formation.[45]
Niche partitioning has been reported among Bayan Shireh species, such is the case of high browser therizinosaurids Erlikosaurus and Segnosaurus,[36] or the grazer Talarurus and browser Tsagantegia.[39] Erketu may have been the tallest herbivore of the paleofauna.[7]
See also
References
- Perle, A.; Norell, M. A.; Clark, J. M. (1999). "A new maniraptoran Theropod−Achillobator giganticus (Dromaeosauridae)−from the Upper Cretaceous of Burkhant, Mongolia". Contributions from the Geology and Mineralogy Chair, National Museum of Mongolia (101): 1−105. OCLC 69865262.
- Senter, P. (2007). "A method for distinguishing dromaeosaurid manual unguals from pedal sickle claws" (PDF). Bulletin of Gunma Museum of Natural History (11): 1–6. ISSN 1342-4092.
- Turner, A. H.; Makovicky, P. J.; Norell, M. A. (2012). "A Review of Dromaeosaurid Systematics and Paravian Phylogeny". Bulletin of the American Museum of Natural History (371): 1−206. doi:10.1206/748.1. hdl:2246/6352. S2CID 83572446.
- Burnham, D. A.; Derstler, K. L.; Currie, P. J.; Bakker, R. T.; Zhou, Z.; Ostrom, J. H. (2000). "Remarkable New Birdlike Dinosaur (Theropoda: Maniraptora) from the Upper Cretaceous of Montana". The University of Kansas Paleontological Contributions. 13: 1−14. doi:10.17161/PCNS.1808.3761. hdl:1808/3761. ISSN 1046-8390.
- Norell, M. A.; Makovicky, P. J. (2004). "Dromaeosauridae". In Weishampel, D. B.; Dodson, P.; Osmólska, H. (eds.). The Dinosauria (2nd ed.). University of California Press. p. 207. ISBN 9780520941434.
- Turner, A.H.; Pol, D.; Clarke, J.A.; Erickson, G.M.; Norell, M.A. (2007). "A Basal Dromaeosaurid and Size Evolution Preceding Avian Flight". Science. 317 (5843): 1378−1381. Bibcode:2007Sci...317.1378T. doi:10.1126/science.1144066. PMID 17823350. Supporting Online Material
- Paul, G. S. (2016). The Princeton Field Guide to Dinosaurs (2nd ed.). Princeton, New Jersey: Princeton University Press. p. 151. ISBN 9780691167664.
- Ely, R. C.; Case, J. A. (2019). "Phylogeny of a new gigantic paravian (Theropoda; Coelurosauria; Maniraptora) from the Upper Cretaceous of James Ross Island, Antarctica". Cretaceous Research. 101: 1−16. doi:10.1016/j.cretres.2019.04.003.
- Turner, A.H.; Makovicky, P.J.; Norell, M.A. (2007). "Feather quill knobs in the dinosaur Velociraptor". Science. 317 (5845): 1721. Bibcode:2007Sci...317.1721T. doi:10.1126/science.1145076. PMID 17885130.
- Lü, J.; Brusatte, S.L. (2015). "A large, short-armed, winged dromaeosaurid (Dinosauria: Theropoda) from the Early Cretaceous of China and its implications for feather evolution". Scientific Reports. 11775 (5): 11775. Bibcode:2015NatSR...511775L. doi:10.1038/srep11775. PMC 4504142. PMID 26181289.
- Longrich, N. R.; Currie, P. J. (2009). "A microraptorine (Dinosauria–Dromaeosauridae) from the Late Cretaceous of North America". Proceedings of the National Academy of Sciences. 106 (13): 5002−5007. Bibcode:2009PNAS..106.5002L. doi:10.1073/pnas.0811664106. PMC 2664043. PMID 19289829.
- Han, G.; Chiappe, L. M.; Ji, S.-A.; Habib, M.; Turner, A. H.; Chinsamy, A.; Liu, X.; Han, L. (2014). "A new raptorial dinosaur with exceptionally long feathering provides insights into dromaeosaurid flight performance". Nature Communications. 5 (4382): 4382. Bibcode:2014NatCo...5.4382H. doi:10.1038/ncomms5382. PMID 25025742.
- Cau, A.; Beyrand, V.; Voeten, D. F. A. E.; Fernandez, V.; Tafforeau, P.; Stein, K.; Barsbold, R.; Tsogtbaatar, K.; Currie, P. J.; Godefroit, P. (2017). "Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs". Nature. 552 (7685): 395–399. Bibcode:2017Natur.552..395C. doi:10.1038/nature24679. PMID 29211712. S2CID 4471941.
- Hartman, S.; Mortimer, M.; Wahl, W.R.; Lomax, D.R.; Lippincott, J.; Lovelace, D.M. (2019). "A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight". PeerJ. 7: e7247. doi:10.7717/peerj.7247. PMC 6626525. PMID 31333906.
- Currie, P. J.; Evans, D. C. (2020). "Cranial Anatomy of New Specimens of Saurornitholestes langstoni (Dinosauria, Theropoda, Dromaeosauridae) from the Dinosaur Park Formation (Campanian) of Alberta". The Anatomical Record. 303 (4): 691–715. doi:10.1002/ar.24241. PMID 31497925. S2CID 202002676.
- Senter, P.; Barsbold, R.; Britt, B. B; Burnham, D. A. (2004). "Systematics and evolution of Dromaeosauridae (Dinosauria, Theropoda)". Bulletin of Gunma Natural History Museum. 8: 1–20.
- Senter, P.; Kirkland, J. I.; Deblieux, D. D.; Madsen, S.; Toth, N. (2012). "New Dromaeosaurids (Dinosauria: Theropoda) from the Lower Cretaceous of Utah, and the Evolution of the Dromaeosaurid Tail". PLOS ONE. 7 (5): e36790. Bibcode:2012PLoSO...736790S. doi:10.1371/journal.pone.0036790. PMC 3352940. PMID 22615813.
- Britt, B. B.; Chure, D. J.; Stadtman, K. L.; Madsen, J. H.; Scheetz, R. D.; Burge, D. L. (2001). "New osteological data and the affinities of Utahraptor from the Cedar Mountain Fm. (Early Cretaceous) of Utah". Journal of Vertebrate Paleontology. 21 (supp. 3): 36A. doi:10.1080/02724634.2001.10010852. S2CID 220414868.
- Jasinski, S. E.; Sullivan, R. M.; Dodson, P. (2020). "New Dromaeosaurid Dinosaur (Theropoda, Dromaeosauridae) from New Mexico and Biodiversity of Dromaeosaurids at the end of the Cretaceous". Scientific Reports. 10 (1): 5105. Bibcode:2020NatSR..10.5105J. doi:10.1038/s41598-020-61480-7. ISSN 2045-2322. PMC 7099077. PMID 32218481.
- Scott Persons IV, W.; Currie, P. J. (2016). "An approach to scoring cursorial limb proportions in carnivorous dinosaurs and an attempt to account for allometry". Scientific Reports. 6 (19828): 19828. Bibcode:2016NatSR...619828P. doi:10.1038/srep19828. PMC 4728391. PMID 26813782.
- Manning, P.L.; Margetts, L.; Johnson, M.R.; Withers, P.J.; Sellers, W.I.; Falkingham, P.L.; Mummery, P.M.; Barrett, P.M.; Raymont, D.R. (2009). "Biomechanics of dromaeosaurid dinosaur claws: Application of X-ray microtomography, nanoindentation, and finite element analysis". The Anatomical Record. 292 (9): 1397–1405. doi:10.1002/ar.20986. PMID 19711472. S2CID 12902686.
- "Killing claws of Velociraptor… are for climbing". New Scientist. 203 (2725): 10. 2009. doi:10.1016/s0262-4079(09)62381-2.
- Fowler, D.W.; Freedman, E.A.; Scannella, J.B.; Kambic, R.E. (2011). "The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds". PLOS ONE. 6 (12): e28964. Bibcode:2011PLoSO...628964F. doi:10.1371/journal.pone.0028964. PMC 3237572. PMID 22194962.
- Powers, M. A.; Sullivan, C.; Currie, P. J. (2020). "Re-examining ratio based premaxillary and maxillary characters in Eudromaeosauria (Dinosauria: Theropoda): Divergent trends in snout morphology between Asian and North American taxa". Palaeogeography, Palaeoclimatology, Palaeoecology. 547 (109704): 109704. Bibcode:2020PPP...547j9704P. doi:10.1016/j.palaeo.2020.109704.
- Khand, Y.; Badamgarav, D.; Ariunchimeg, Y.; Barsbold, R. (2000). "Cretaceous system in Mongolia and its depositional environments". Cretaceous Environments of Asia. Developments in Palaeontology and Stratigraphy. 17. pp. 49–79. doi:10.1016/s0920-5446(00)80024-2. ISBN 9780444502766.
- Shuvalov, V.F. (2003). "The Cretaceous stratigraphy and palaeobiogeography of Mongolia". The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press. pp. 256–278. ISBN 9780521545822.
- Eberth, D.A.; Kobayashi, Y.; Lee, Y.N.; Mateus, O.; Therrien, F.; Zelenitsky, D.K.; Norell, M.A. (2009). "Assignment of Yamaceratops dorngobiensis and associated redbeds at Shine Us Khudag (eastern Gobi, Dorngobi Province, Mongolia) to the redescribed Javkhlant Formation (Upper Cretaceous)". Journal of Vertebrate Paleontology. 29 (1): 295–302. doi:10.1080/02724634.2009.10010384. S2CID 197540125.
- Averianov, A.; Sues, H. (2012). "Correlation of Late Cretaceous continental vertebrate assemblages in Middle and Central Asia" (PDF). Journal of Stratigraphy. 36 (2): 462–485. S2CID 54210424.
- Tsogtbaatar, K.; Weishampel, D.B.; Evans, D.C.; Watabe, M. (2019). "A new hadrosauroid (Dinosauria: Ornithopoda) from the Late Cretaceous Baynshire Formation of the Gobi Desert (Mongolia)". PLOS ONE. 14 (4): e0208480. Bibcode:2019PLoSO..1408480T. doi:10.1371/journal.pone.0208480. PMC 6469754. PMID 30995236.
- Hicks, J.F.; Brinkman, D.L.; Nichols, D.J.; Watabe, M. (1999). "Paleomagnetic and palynologic analyses of Albian to Santonian strata at Bayn Shireh, Burkhant, and Khuren Dukh, eastern Gobi Desert, Mongolia". Cretaceous Research. 20 (6): 829–850. doi:10.1006/cres.1999.0188.
- Kurumada, Y.; Aoki, S.; Aoki, K.; Kato, D.; Saneyoshi, M.; Tsogtbaatar, K.; Windley, B. F.; Ishigaki, S. (2020). "Calcite U–Pb age of the Cretaceous vertebrate‐bearing Bayn Shire Formation in the Eastern Gobi Desert of Mongolia: usefulness of caliche for age determination". Terra Nova. 32 (4): 246–252. doi:10.1111/ter.12456.
- Jerzykiewicz, T.; Russell, D. A. (1991). "Late Mesozoic stratigraphy and vertebrates of the Gobi Basin". Cretaceous Research. 12 (4): 345−377. doi:10.1016/0195-6671(91)90015-5.
- Tsuihiji, T.; Watabe, M.; Barsbold, R.; Tsogtbaatar, K. (2015). "A gigantic caenagnathid oviraptorosaurian (Dinosauria: Theropoda) from the Upper Cretaceous of the Gobi Desert, Mongolia". Cretaceous Research. 56: 60−65. doi:10.1016/j.cretres.2015.03.007.
- Guo, Z. X.; Shi, Y. P.; Yang, Y. T.; Jiang, S. Q.; Li, L. B.; Zhao, Z. G. (2018). "Inversion of the Erlian Basin (NE China) in the early Late Cretaceous: Implications for the collision of the Okhotomorsk Block with East Asia" (PDF). Journal of Asian Earth Sciences. 154: 49–66. Bibcode:2018JAESc.154...49G. doi:10.1016/j.jseaes.2017.12.007.
- Ksepka, D.T.; Norell, M.A. (2006). "Erketu ellisoni, a long-necked sauropod from Bor Guvé (Dornogov Aimag, Mongolia)" (PDF). American Museum Novitates. 2006 (3508): 1–16. doi:10.1206/0003-0082(2006)3508[1:EEALSF]2.0.CO;2.
- Zanno, L. E.; Tsogtbaatar, K.; Chinzorig, T.; Gates, T. A. (2016). "Specializations of the mandibular anatomy and dentition of Segnosaurus galbinensis (Theropoda: Therizinosauria)". PeerJ. 4: e1885. doi:10.7717/peerj.1885. PMC 4824891. PMID 27069815.
- Kobayashi, Y.; Barsbold, R. (2005). "Reexamination of a primitive ornithomimosaur, Garudimimus brevipes Barsbold, 1981 (Dinosauria: Theropoda), from the Late Cretaceous of Mongolia" (PDF). Canadian Journal of Earth Sciences. 42 (9): 1501−1521. Bibcode:2005CaJES..42.1501K. doi:10.1139/e05-044. hdl:2115/14579.
- Perle, A. (1977). "O pervoy nakhodke Alektrozavra (Tyrannosauridae, Theropoda) iz pozdnego Mela Mongolii" [On the first discovery of Alectrosaurus (Tyrannosauridae, Theropoda) in the Late Cretaceous of Mongolia]. Shinzhlekh Ukhaany Akademi Geologiin Khureelen (in Russian). 3 (3): 104–113.
- Park, J. Y.; Lee, Y. N.; Currie, P. J.; Kobayashi, Y.; Koppelhus, E.; Barsbold, R.; Mateus, O.; Lee, S.; Kim, S. H. (2020). "Additional skulls of Talarurus plicatospineus (Dinosauria: Ankylosauridae) and implications for paleobiogeography and paleoecology of armored dinosaurs" (PDF). Cretaceous Research. 108 (104340). doi:10.1016/j.cretres.2019.104340.
- Watabe, M.; Tsogtbaatar, K.; Sullivan, R.M. (2011). "A new pachycephalosaurid from the Baynshire Formation (Cenomanian-late Santonian), Gobi Desert, Mongolia" (PDF). Fossil Record 3. New Mexico Museum of Natural History and Science, Bulletin. 53: 489–497.
- Sereno, P.C. (2000). "The fossil record, systematics and evolution of pachycephalosaurs and ceratopsians from Asia" (PDF). The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press. pp. 489–491.
- Sukhanov, V.B.; Danilov, I.G.; Syromyatnikova, E.V. (2008). "The Description and Phylogenetic Position of a New Nanhsiungchelyid Turtle from the Late Cretaceous of Mongolia". Acta Palaeontologica Polonica. 53 (4): 601–614. doi:10.4202/app.2008.0405.
- Danilov, I.G.; Hirayama, R.; Sukhanov, V.B.; Suzuki, S.; Watabe, M.; Vitek, N.S. (2014). "Cretaceous soft-shelled turtles (Trionychidae) of Mongolia: new diversity, records and a revision". Journal of Systematic Palaeontology. 12 (7): 799–832. doi:10.1080/14772019.2013.847870. S2CID 86304259.
- Turner, A.H. (2015). "A Review of Shamosuchus and Paralligator (Crocodyliformes, Neosuchia) from the Cretaceous of Asia". PLOS ONE. 10 (2): e0118116. Bibcode:2015PLoSO..1018116T. doi:10.1371/journal.pone.0118116. PMC 4340866. PMID 25714338.
- Rougier, G. W.; Davis, B. M.; Novacek, M. J. (2015). "A deltatheroidan mammal from the Upper Cretaceous Baynshiree Formation, eastern Mongolia". Cretaceous Research. 52: 167−177. doi:10.1016/j.cretres.2014.09.009.
External links
 Media related to Achillobator at Wikimedia Commons
Media related to Achillobator at Wikimedia Commons Data related to Achillobator at Wikispecies
Data related to Achillobator at Wikispecies





.png.webp)



.jpg.webp)





