Therizinosaurus
Therizinosaurus (/ˌθɛrɪˌzɪnoʊˈsɔːrəs/; meaning "scythe lizard") is a genus of very large therizinosaurid that lived in Asia during the Late Cretaceous period in what is now the Nemegt Formation around 70 million years ago. The first remains of Therizinosaurus were found in 1948 by a Mongolian field expedition at the Gobi Desert, and later described by Evgeny Maleev in 1954 who originally thought the remains to belong to a turtle-like reptile. Therizinosaurus is known only from a few bones, including gigantic hand claws, from which it gets its name, and additional findings comprising forelimb and hindlimb elements that have been discovered during the 1960s-1980s.
| Therizinosaurus | |
|---|---|
 | |
| Mounted forelimbs of specimen MPC-D 100/15 at Nagoya City Science Museum | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Superfamily: | †Therizinosauroidea |
| Family: | †Therizinosauridae |
| Genus: | †Therizinosaurus Maleev, 1954 |
| Type species | |
| †Therizinosaurus cheloniformis Maleev, 1954 | |
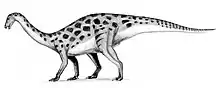
Therizinosaurus comprises the single species T. cheloniformis, which could grow up from 9 to 10 m (30 to 33 ft) long and weigh possibly over 3 t (3,000 kg). It had the longest known claws of any land animal, reaching above 50 cm (20 in) in length. Unlike other therizinosaurs, the claws were very stiff and elongated, but like other members, it would have been a slow-moving, long-necked/high browser herbivore equipped with a rhamphotheca (horny beak) and a wide torso for food processing. The feet ended in four weight-bearing toes, resembling the unrelated sauropodomorphs.
It was one of the last and the largest representative of its unique group, the Therizinosauria. Therizinosaurus is classified as a therizinosaurid, meaning it was more derived (advanced) than the primitive therizinosauroids. At first, therizinosaurs had very complex relationships due to the lack of genera and findings at the time. Originally, they were thought to be some kind of Cretaceous sauropodomorphs or transitional ornithischians, but after years of taxonomic debate they are now placed within the Theropoda, specifically as maniraptorans. The unusually elongated hand claws of Therizinosaurus were more useful when pulling vegetation within reach rather than being used for attack or defense. Therizinosaurus was a very tall animal, likely having a reduced competition over the foliage and outmatching predators like Tarbosaurus.
History of discovery
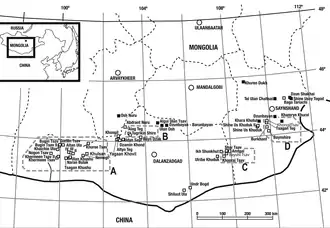
In 1948, several Mongolian Paleontological expeditions organized by the USSR Academy of Sciences were conducted in the Nemegt Formation of the Gobi Desert, Southwestern Mongolia, with the main objective of new fossils findings. The expeditions unearthed numerous dinosaur and turtle fossil remains from the stratotype locality Nemegt (also known as Nemegt Valley), but the most notable elements collected were three partial manual unguals (claw bones) of considerable size. This set of unguals was found on a subdivision of the Nemegt locality designated as the fifth quarry near the skeleton of a large theropod, but also in association with other elements including a metacarpal fragment and several ribs fragments. It was labelled under the specimen number PIN 551-483 and later on, these fossils were described by the Russian paleontologist Evgeny Maleev in 1954 who used them to scientifically name the new genus and type species Therizinosaurus cheloniformis, becoming the holotype specimen. The generic name, Therizinosaurus, is derived from the Greek θερίζω (therízo, meaning scythe, reap or cut) and σαῦρος (sauros, meaning lizard) in reference to the enormous manual unguals, and the specific name, cheloniformis, is taken from the Greek χελώνη (chelóni, meaning turtle) and Latin formis as the remains were thought to belong to a turtle-like reptile. Maleev also coined a separate family for this new and enigmatic taxon: Therizinosauridae. Since little was known of Therizinosaurus at the time of the original description, Maleev thought PIN 551-483 belonged to a large, 4.5 m (15 ft) long marine turtle that relied on its giant hand claws to harvest seaweed.[1]

Though it was not fully understood what general kind of animal these fossils belonged, in 1970 the Russian paleontologist Anatoly K. Rozhdestvensky was one of the first authors in suggest that Therizinosaurus was a theropod and not a turtle. He made comparisons between Chilantaisaurus and the holotype unguals of Therizinosaurus to propose that these appendages actually came from a carnosaurian dinosaur, thereby interpreting Therizinosaurus as a theropod. Rozhdestvensky also illustrated the three holotypic manual unguals and re-identified the metacarpal fragment as a metatarsal bone, and based on the unusual shape of both metatarsal and ribs fragments he listed them as sauropod remains.[2] These theropodan affinities were also followed by the Polish paleontologist Halszka Osmólska and co-author Ewa Roniewicz in 1970 during their naming and description of Deinocheirus—another large and enigmatic theropod from the formation that was initially known from partial arms. Similar to Rozhdestvensky, they suggested that the holotype unguals were more likely to have belonged to a carnosaurian theropod, rather than to a large marine turtle.[3]
Additional specimens

Further expeditions in the Nemegt Formation unearthed more fossils of Therizinosaurus. In 1968 prior to Rozhdestvensky, Osmólska and Roniewicz statements, the upper portion of a manual ungual was found in the Altan Uul locality and labelled as MPC-D 100/17 (formerly IGM or GIN). In 1972, another fragmented ungual (specimen MPC-D 100/16) was discovered at the Upper White Beds of the Hermiin Tsav locality, only preserving its lower portion.[4][5] During the year 1973, a much complete, larger, and articulated specimen was collected also from the Nemegt locality. This specimen was labelled is MPC-D 100/15 and consists of both left and right arms including the scapulocoracoids, both humeri (upper arm bones), right ulna with radius and left ulna, two right carpals, the right metacarpus including a complete digit Il, and some ribs with gastralia (belly ribs). As common with fossils, some elements were not entirely preserved such as the scapulocoracoids with broken ends, and the left arm is less complete than the right one. All of these specimens were first described and referred to Therizinosaurus by the Mongolian paleontologist Rinchen Barsbold in 1976. In this new monograph, he pointed out that the rib fragments in MPC-D 100/15 were more slender than the ones from the holotype, and identified MPC-D 100/16 and 100/17 as pertaining to digits I and III, respectively. It was clear to Barsbold that MPC-D 100/15 represented Therizinosaurus as the ungual in this specimen shared the elongation and flattened morphology of all previous specimens. He concluded that Therizinosaurus was a theropod taxon since MPC-D 100/15 matched multiple theropodan characters.[4]
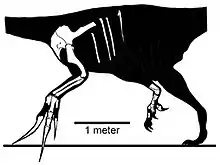
Also during the year 1973, the specimen MPC-D 100/45 was discovered by the Joint Soviet-Mongolian Paleontological Expedition at the Hermiin Tsav locality. Unlike the previous findings, MPC-D 100/45 is represented by a right hindlimb composed of a very fragmented femur with the lower end of the tibia, astragalus, calcaneum, tarsal IV, a functional tetradacyl feet (four-toed) compromising four partial metatarsals, partially preserved digits I and III, and nearly complete digits II and IV. These newer remains were described by the also Mongolian paleontologist Altangerel Perle in 1982. He also regarded the referral of Therizinosaurus and Therizinosauridae to Chelonia (turtles order) to be unlikely, and hypothesized Therizinosaurus and Segnosaurus—at the time of this description regarded as a theropod dinosaur—to be particularly similar based on their respective scapulocoracoid morphology, only differing in size. Perle referred MPC-D 100/45 to Therizinosaurus given that this specimen was found near the location of MPC-D 100/15 and virtually similar to the described pes for Segnosaurus.[6] In 1990, Barsbold and Teresa Maryanska agreed with Perle in that the hindlimb material from Hermiin Tsav he described in 1982 was therizinosaurian (then called segnosaurians) given that the metatarsus was stocky and the astragalus had a laterally arched ascending process (bony extension), but cast doubt with his referral of it to Therizinosaurus and the segnosaurian identity for this taxon since it was only known from the pectoral girdle and other forelimb elements, making direct comparisons between specimens impossible. They considered this specimen to represent a Late Cretaceous representative of the Segnosauria, but not Therizinosaurus.[7]
In 2010 however, the North American paleontologist Lindsay E. Zanno in her large taxonomic reevaluation of Therizinosauria considered the referral of MPC-D 100/45 to Therizinosaurus to be likely based on the rationale that it was collected in the same stratigraphic context (Nemegt Formation) as the holotype, and shared the robust and four-toed morphology of other therizinosaurids such as Segnosaurus. She also excluded the rib material from the holotype as it was re-identified by Rozhdestvensky to likely have come from a sauropod dinosaur, and not Therizinosaurus itself.[8]
Description
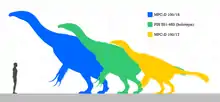
For maniraptoran standards, Therizinosaurus obtained enormous sizes, estimated to have reach 9 to 10 m (30 to 33 ft) in length with ponderous weights from 3 t (3,000 kg) to possibly over 5 t (5,000 kg).[9][10][11] These dimensions make Therizinosaurus the largest therizinosaur known and the largest known maniraptoran. Along with the contemporaneous Deinocheirus, it was the largest maniraptoriform.[12] Though the body remains of Therizinosaurus are relatively incomplete, inferences can be made about its physical characteristics based on more complete and related therizinosaurids. Like other members of their family, Therizinosaurus had a proportionally small skull bearing a rhamphotheca (horny beak) atop the long neck, with bipedal gaits and a heavy, broad belly for foliage processing with the addition of sparse feathering. Its hindlimbs ended in four functionally, weight-bearing toes unlike other theropod groups in which the first toe was reduced to a dewclaw.[13][14][15] In 2010, Senter and James used hindlimb length equations to predict the total length of the hindlimbs in Therizinosaurus and Deinocheirus. They concluded that an average Therizinosaurus may have had approximately 3 m (9.8 ft) long legs.[16] More recently, Mike Taylor and Matt Wedel suggested that the whole neck would be 2.9 times the size of the humerus, which was 76 cm (760 mm), resulting in a 2.2 m (7.2 ft) long neck based on comparisons with the cervical vertebrae series of Nanshiungosaurus.[17]

Therizinosaurus can be distinguished from other therizinosaurids in having very straight manual unguals that are side to side compressed (flattened) and hypertrophied (elongated) with poor curves, an extended deltopectoral crest on the humerus, the length of metacarpal I is larger than 2/3 the length of metacarpal III, and the metacarpal I has an enlarged inner crest that connects the inner lower condyle and upper inner lobe.[8] The most distinctive feature of Therizinosaurus was the presence of gigantic claws on each of the three digits of its front limbs. These were common among therizinosaurs but particularly large and stiffened in Therizinosaurus, and they are considered as the longest known from any terrestrial animal.[12]
Forelimbs

The most complete arm of Therizinosaurus is represented by the specimen MPC-D 100/15 which preserves an almost complete right arm and a more fragmented left arm. The right arm covers 2.4 m (7.9 ft) in total length.[16] The scapula measures 67 cm (670 mm) long with a stocky and flattened dorsal blade, wide acromial process and a very widened ventral surface; the blade seems to be broken at the end though. Near the anterior edge of the scapular widening and near the scapulocoracoid suture, a foramen is visible; it likely functioned as a channel for blood vessels and nerves when alive. The posterior edge of the scapula is robust and the acromion is lightly built, likely fused into a cartilaginous system with its periphery in life. The coracoid measures 36 cm (360 mm) in length, it has a broad and convex lateral surface that forms a slightly inclined concavity near of the scapulocoracoid suture. This concavity bends down towards the scapular widening. Near the scapulocoracoid suture, this edge turns very thin and possibly into cartilage along with the periphery of the coracoid in life, as the case of the scapular edge. A large foramen is also present on the coracoid. The glenoid is broad and deep, slightly pointing to the outer lateral side. It has robust, convex crest-like borders. The supraglenoid thickness is developed in a convex crest-shaped form, it is divided across the top of the scapulocoracoid suture. The attachment for the biceps muscle is prominently developed by a large tubercle with a stocky top, indicating powerful muscles in life.[18]
The humerus is robustly built, measuring 76 cm (760 mm) long. It has a broad proximal end. The deltoid crest is tall and thick, its top is located approximately 1/3 from the upper end, the length of the crest is no less than 2/3 the length of the whole bone element. The lower end of the humerus is very expanded and flattened in a posterior direction. The condyles are developed onto the anterior side of the lower expansion while the epicondyles are very broad and project over the limits of the articular areas. The ulna is measured at 62.02 cm (620.2 mm) and most of its length is occupied by the straight shaft. The ulnar process is very wide. The upper articular area can be divided into the inner and outer lateral sides. The lateral side has a triangular-shaped border and is slightly concave; it is limited in a top view by the fossa for the upper articulation of the radius. The inner side forms a semilunar-shaped fossa that caps the lunar-shaped condyle of the humerus. The radius is 55.04 cm (550.4 mm) long and slightly S-curved. The upper end is flattened in a lateral direction, very wide and the distal end is highly robust.[18]
Hand and claws

The preserved right manus is composed of two lower carpal bones, complete three metacarpal bones and a complete second digit. The first lower carpal measures 8.23 cm (82.3 mm) tall and 8.53 cm (85.3 mm) wide having two articulations on the lowermost end. The upper surface of this carpal is divided by a broad fossa that forms the articulation of the carpus. On its inner side it has a triangular-shaped outline and attaches to the upper surface of metacarpal I occupying a little bit less than the lateral side, which articulates to metacarpal II. When they cross, these areas are separated by an oblique projection. The second lower carpal is smaller than the first one, it measures 5.6 cm (56 mm) tall and 5.93 cm (59.3 mm) tall. Its lower surface is flattened and the articular surface of the carpus extends from the first carpal to the second carpal over the articulation of the two bones.[18]
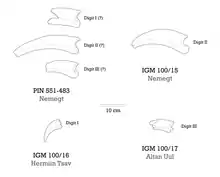
The metacarpal I is 14.55 cm (145.5 mm) long and compared to the others is more robust and stocky. Overall, the lateral side is broad, especially on the uppermost area; the inner border is thin and narrow. The upper articulation is configured into three parts. The lower articular top is somewhat asymmetric, and bends to the inner side from the left, along with a wide, deep opening. The metacarpal II measures 28.68 cm (286.8 mm) in length and is the most elongated and robust. It has an inclined, square and flattened upper articulation. The articulation on the lower head has very symmetrical condyles, being divided by a broad, deep fossa. The lateral connecting openings are poorly developed. The metacarpal III covers 19.16 cm (191.6 mm) in length, it has a very thin shaft compared to the other metacarpals, and features an upper articulation that splits into three parts. The lower articular head is asymmetrical with deep and broad openings. As in metacarpal II, the lateral connecting openings are poorly developed. Only the second digit of the right manus was preserved, consisting of two phalanges and a large ungual. The first and second phalanges are somewhat equal in shape and length (14.17 cm (141.7 mm) and 14.38 cm (143.8 mm), respectively), they also share the robust and stocky structure. The upper articular facets are very symmetrical and preserve a crest that is particularly taller in the first phalanx. The top border of this crest is very pointed and thick; it likely served as the site for attachment of the extensor tendons in life. The lower heads are nearly symmetrical, but the central fossa is considerably wider and deeper in the first phalanx. The preserved unguals are specially enormous, estimated to be approximately 52 cm (520 mm) in length. Unlike other therizinosaurs, they are very straight, laterally flattened and not particularly curved with very sharp point tips. The upper articulation is symmetrical and has a cross-ridge that divides the surface into two asymmetrical parts. The size of the unguals MPC-D 100/16 and 100/17 may indicate that they belong to manual digits I and III, respectively.[18]
Classification
Maleev originally classified Therizinosaurus as to have been a giant marine turtle and the genus was assigned by Maleev to a separate Therizinosauridae given how enigmatic the specimen was.[1] The fossils remained with uncertainty among the scientific community, however, in 1970 Rozhdestvensky was one of the first paleontologists suggesting that Therizinosaurus was actually a theropod dinosaur instead of some turtle. Also, the supposed ribs of the holotype were likely from a different dinosaur, possibly some sauropodomorph.[2] With the discovery and description of Segnosaurus, in 1979 Perle named a new family of dinosaurs, the Segnosauridae. He tentatively placed the family within Theropoda given the similarities of the mandible and dentition to other members.[19] A year later, the new genus Erlikosaurus was named by Barsbold and Perle in 1980. They named a new infraorder called the Segnosauria, composed by Erlikosaurus and Segnosaurus. They also noted that while aberrant and having ornithischian-like pelves, segnosaurs featured similar traits to other theropods.[20] With the discovery of the referred hindlimb to Therizinosaurus in 1982 by Perle, he concluded that Segnosaurus was very similar to the latter based on the pes morphology and they possibly belonged to a single, if not the same group.[6] In 1983, Barsbold named a new genus of segnosaur Enigmosaurus. He analysed the pelvis of this new genus and pointed out that segnosaurids were so different from other theropods that they could be outside the group or represent a different lineage of theropod dinosaurs.[18] Later on the same year, he intensified the exclusion of segnosaurs from being theropods by noting that their pelves resembled those of sauropod dinosaurs.[21]

Consequently, the assignment of segnosaurs started to shift towards sauropodomorphs. In 1984, Paul claimed that segnosaurs rather than being aberrant theropods were indeed, sauropodomorphs which successfully managed to remain into the Cretaceous period. He based this idea on anatomical traits such as the skull and similar pes configuration.[22] He maintained his position in 1988 by placing the Segnosauria into the now obsolete Phytodinosauria, and was one of the first to suggest a segnosaur assignment for the enigmatic Therizinosaurus.[23] Other prominent paleontologists like Gauthier or Sereno supported this vision.[24][25] In 1990, Barsbold and Teresa Maryanska agreed in that the hindlimb material from Hermiin Tsav referred to Therizinosaurus in 1982 was segnosaurian since it matched several traits, but considered it unlikely to belong to the genus and species as there was no overlapping material among specimens. Barsbold and Maryanska also disagreed with previous researchers who classified Deinocheirus as a segnosaur.[7] In the same year, David B. Norman considered Therizinosaurus to be a theropod of uncertain classification.[26]

However, with the unexpected discovery and description of Alxasaurus in 1993, the widely accepted sauropodomorph affinities of segnosaurs were questioned by Rusell and Dong. This new genus was far more complete than any other segnosaur and multiple anatomical features indicated that it was related to Therizinosaurus. With this, they identified the Therizinosauridae along with the Segnosauridae to be the same group, the former name having taxonomic priority. Due to some primitive characters present in Alxasaurus they coined a new taxonomic rank, the Therizinosauroidea, containing this new taxon and the Therizinosauridae. All of this new information provided data on the affinities of the new-named therizinosauroids. Lastly, Russell and Dong concluded that these animals were actual theropods with unusually aberrant features.[27] In 1994, Clark and colleagues redescribed the very complete skull of Erlikosaurus and even more theropod traits were found this time. They also validated the synonymy of the Segnosauridae with Therizinosauridae and considered therizinosauroids as maniraptoran dinosaurs.[28] In 1997, Rusell coined the infraorder Therizinosauria in order to contain all segnosaurs. This new infraorder was composed of Therizinosauroidea and the more advanced Therizinosauridae. With this, Segnosauria became a synonym of Therizinosauria.[29] Though some uncertainties remained, a small and feathered therizinosauroid from China was described in 1999 by Xu and colleagues, the new genus Beipiaosaurus. It definitely confirmed the placement of therizinosaurs among theropods and also their taxonomic place on the Coelurosauria. This discovery also indicated that feathers were highly distributed among theropod dinosaurs.[30]
In 2010, Lindsay Zanno revised the taxonomy of therizinosaurs with extensive detail. She found that many parts on therizinosaur holotypes were lost or damaged, and sparse specimens with no overlapping elements were disadvantages when concluding the relationships of the members. Zanno accepted the referral of the specimen IGM 100/45 to Therizinosaurus since it matches multiple therizinosaurid traits but decided not include the specimen in her taxonomic analysis due to the lack of forelimb remains. She also excluded the supposed ribs that were present on the holotype since they likely came from a different animal and not Therizinosaurus.[8] In 2019, Hartman and colleagues also performed a large phylogenetic analysis of Therizinosauria based on the characters provided by Zanno in her revision. They found similar results to Zanno regarding the family Therizinosauridae but this time with the inclusion of more taxa and specimens. Cladogram below shows the placement of Therizinosaurus within Therizinosauria according to Hartman and colleagues in 2019:[31]
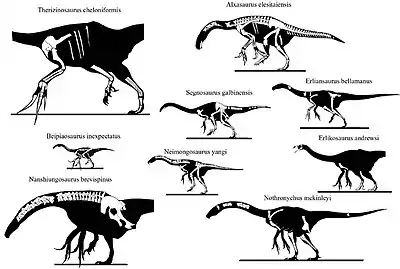
| Therizinosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiology
In 2012, Stephan Lautenschlager and colleagues analyzed the complete endocranial anatomy of Erlikosaurus and other therizinosaurs that preserve braincases. They found that therizinosaurids had well-developed senses of smell, hearing, and balance. The former two senses may have played an important role during foraging, predator evasion, and complex social behavior. These senses were also well-developed in earlier coelurosaurs and other theropods, indicating that therizinosaurs may have inherited many of these traits from their carnivorous ancestors and used them for their different and specialized dietary purposes.[32]
The discovery of fossilized embryos at the Nanchao Formation and multiple egg nests from other formations indicates that therizinosaurs had a colonial nesting-style similar to other herbivores like some sauropodomorphs, titanosaurs and hadrosaurs. Nestling therizinosaurs were capable of movements after birth and did not necessarily depend of their parents.[33] It is also indicated that therizinosaurs did not have a philopatric behaviour when nesting.[34][35]
Claws function

When first described by Maleev in 1954, he considered that the unusually large claws were used to harvest seaweed. This however, based on the assumption of a giant marine turtle.[1] In 1970, Rozhdestvensky re-examined the claws and suggested a possible function specialized in opening termite mounds.[2] In 1995, Lev A. Nessov suggested the elongated claws were used for defense against predators and juveniles could have used their claws for arboreal locomotion.[36]
In 2014, Lautenschlager tested the function of various therizinosaur hand claws—including Therizinosaurus—through digital simulations. Three different functional scenarios were simulated for each claw morphology with a force of 400 N applied in each scenario: scratch/digging; hook-and-pull; and piercing. Though the stocky claws of Alxasaurus resulted on low the stress magnitudes, the stress was greater with the curvature and elongation of the claws in Falcarius, Nothronychus and Therizinosaurus. Some of the highest stress, deformation and strain magnitudes were obtained in the scratch/digging scenario, the hook-and-pull scenario in contrast, resulted in lower magnitudes, and lesser ones were found in the piercing scenario. Particularly, the overall stress was most pronounced in the unusual claws of Therizinosaurus, which may represent an exceptional case of elongation specialization. Lautenschlager noted the more strongly curved and elongate claws of some therizinosaurian taxa were poorly functional in a scratch/digging fashion, indicating this as the most unlikely function. Though fossorial behaviour has been reported in several dinosaur species, the large body size largely rules out the possibility of burrow digging in therizinosaurs. Nevertheless, an overall digging action would have been done with the foot claws because, since as in other maniraptorans, feathers on the arms would have interfered with this function. Instead of being used for fossorial behaviour, it is more likely that Therizinosaurus make use of its hands in a hook-and-pull fashion to pull or grasp vegetation within reach. Lautenschlager could neither confirm nor disregard that the hand claws could have been used for defense, intraspecific competition, stabilization by grasping tree trunks during high browsing, sexual dimorphism, or gripping mates during mating given the lack of more specimens. He clarified that there is no evidence that the long claws of Therizinosaurus would have been used in active defense or attack, however, it is possible that these appendages could have had some role when facing a threat.[37]
Paleoenvironment
.jpg.webp)
The remains of Therizinosaurus were found in the well-known Nemegt Formation in the Gobi Desert, dating back to the Maastrichtian stage about 70 million and 68 million years ago.[38] The environments that Therizinosaurus inhabited have been determined by the δ13C level preserved on the tooth enamel of many herbivorous dinosaurs and the numerous petrified wood across the formation, and consisted of large meandering and braided rivers with extensive woodlands composed of large, enclosed, canopy-like forests of Araucarias that supported diverse herbivorous dinosaurs like Therizinosaurus. The climate of the formation was relatively temperate (mean annual temperatures between 7.6-8.7 °C), characterized by monsoons with cold, dry winters and hot, rainy summers with the addition of mean annual precipitations between 775 mmm and 835 mmm, a precipitation that was subject to prominent seasonal fluctuations.[39][38] The wet environments of the Nemegt Formation may have acted as an oasis-like area that attracted oviraptorids from arid neighbour localities such as the Barun Goyot Formation, as evidenced on the presence of Nemegtomaia in both regions.[40]

The dinosaur fauna of the Nemegt Formation was very diverse. Therizinosaurus shared its habitat with other theropods such as the deinonychosaurs Adasaurus and Zanabazar; ornithomimosaurs Anserimimus, Deinocheirus and Gallimimus; oviraptorosaurs Rinchenia, Elmisaurus, Gobiraptor, and the apex predator of the formation, Tarbosaurus. Other herbivorous dinosaurs include Barsboldia, Tarchia, Prenocephale, Nemegtosaurus, Opisthocoelicaudia and the most common herbivore, Saurolophus.[40][41][42]
Senter and James have noted that Therizinosaurus was among the tallest dinosaurs in the Nemegt Formation. Due to its prominent height and high-browsing life-style, it probably had no prominent competition with other herbivores over the foliage. However, they suggested a potential niche partitioning with the sauropods of the formation, which were also long-necked dinosaurs. If engaged in combat, a large Tarbosaurus may have been not able to bite any higher than the thighs or belly of an adult standing Therizinosaurus. The gigantic claws likely functioned as optimal defense against this situation.[16] The paleontologist Thomas Holtz has compared the wet environments of the Nemegt Formation to those of the Okavango Delta. He suggested a resources competition between Deinocheirus, Saurolophus, Therizinosaurus and titanosaurs.[43]
See also
References
- Maleev, E. A. (1954). "Новый черепахообразный ящер в Монголии" [New turtle like reptile in Mongolia]. Priroda (in Russian) (3): 106−108. Translated paper
- Rozhdestvensky, A. K. (1970). "О гигантских когтевых фалангах загадочных рептилий мезозоя" [On the giant claws of enigmatic Mesozoic reptiles]. Paleontological Journal (in Russian). 1970 (1): 131−141.
- Osmólska, H.; Roniewicz, E. (1970). "Deinocheiridae, a new family of theropod dinosaurs" (PDF). Palaeontologica Polonica (21): 5−19.
- Barsbold, R. (1976). "Новые данные о теризинозавре (Therizinosauridae, Theropoda)" [New data on Therizinosaurus (Therizinosauridae, Theropoda)]. In Kramarenko, N. N.; Luvsandansan, B.; Voronin, Y. I.; Barsbold, R.; Rozhdestvensky, A. K.; Trofimov, B. A.; Reshetov, V. Y. (eds.). Paleontology and Biostratigraphy of Mongolia. The Joint Soviet-Mongolian Paleontological Expedition, Transactions (in Russian). Moscow: Nauka Press. pp. 76−92.
- Gradziński, R.; Jaworowska, Z. K.; Maryańska, T. (1977). "Upper Cretaceous Djadokhta, Barun Goyot and Nemegt formations of Mongolia, including remarks on previous subdivisions". Acta Geologica Polonica. 27 (3): 281−326.
- Perle, A. (1982). "On a new finding of the hindlimb of Therizinosaurus sp. from the Late Cretaceous of Mongolia". Problems in Mongolian Geology (in Russian). 5: 94−98. Translated paper
- Barsbold, R.; Maryańska, T. (1990). "Saurischia Sedis Mutabilis: Segnosauria". In Weishampel, D. B.; Osmolska, H.; Dodson, P. (eds.). The Dinosauria (1st ed.). Berkeley: University of California Press. pp. 408−415. ISBN 9780520067271.
- Zanno, L. E. (2010). "A taxonomic and phylogenetic re-evaluation of Therizinosauria (Dinosauria: Maniraptora)". Journal of Systematic Palaeontology. 8 (4): 503−543. doi:10.1080/14772019.2010.488045. S2CID 53405097.
- Holtz, T. R.; Rey, L. V. (2007). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Random House. ISBN 9780375824197. Genus List for Holtz 2012 Weight Information
- Paul, G. S. (2016). The Princeton Field Guide to Dinosaurs (2nd ed.). Princeton, New Jersey: Princeton University Press. pp. 162−168. ISBN 9780691167664.
- Campione, N. E.; Evans, D. C. (2020). "The accuracy and precision of body mass estimation in non‐avian dinosaurs". Biological Reviews. 95 (6): 1759–1797. doi:10.1111/brv.12638. PMID 32869488. S2CID 221404013.
- Molina-Pérez, R.; Larramendi, A. (2016). Récords y curiosidades de los dinosaurios Terópodos y otros dinosauromorfos. Barcelona, Spain: Larousse. p. 270. ISBN 9788416641154.
- Dimond, C. C.; Cabin, R. J.; Brooks, J. S. (2011). "Feathers, Dinosaurs, and Behavioral Cues: Defining the Visual Display Hypothesis for the Adaptive Function of Feathers in Non-Avian Theropods". BIOS. 82 (3): 58–63. doi:10.1893/011.082.0302. JSTOR 23033900. S2CID 98221211.
- Lautenschlager, S.; Lawrence, M. W.; Perle, A.; Zanno, L. E.; Emily, J. R. (2014). "Cranial anatomy of Erlikosaurus andrewsi (Dinosauria, Therizinosauria): new insights based on digital reconstruction". Journal of Vertebrate Paleontology. 34 (6): 1263−1291. doi:10.1080/02724634.2014.874529.
- Hedrick, B. P.; Zanno, L. E.; Wolfe, D. G.; Dodson, P. (2015). "The Slothful Claw: Osteology and Taphonomy of Nothronychus mckinleyi and N. graffami (Dinosauria: Theropoda) and Anatomical Considerations for Derived Therizinosaurids". PLOS ONE. 10 (6): e0129449. Bibcode:2015PLoSO..1029449H. doi:10.1371/journal.pone.0129449. PMC 4465624. PMID 26061728.
- Senter, P.; James, R. H. (2010). "Hip heights of the gigantic theropod dinosaurs Deinocheirus mirificus and Therizinosaurus cheloniformis, and implications for museum mounting and paleoecology" (PDF). Bulletin of Gunma Museum of Natural History (14): 1−10.
- Taylor, M. P.; Wedel, M. J. (2013). "Why sauropods had long necks; and why giraffes have short necks". PeerJ. 1: e36. arXiv:1209.5439. Bibcode:2012arXiv1209.5439T. doi:10.7717/peerj.36. PMC 3628838. PMID 23638372.
- Barsbold, R. (1983). "Хищные динозавры мела Монголии" [Carnivorous dinosaurs from the Cretaceous of Mongolia] (PDF). Transactions of the Joint Soviet-Mongolian Paleontological Expedition (in Russian). 19: 89. Translated paper
- Perle, A. (1979). "Segnosauridae - новое семейство позднемеловых хищных динозавров Монголии" [Segnosauridae - a new family of theropods from the Late Cretaceous of Mongolia]. Transactions of the Joint Soviet-Mongolian Paleontological Expedition (in Russian). 8: 45−55. Translated paper
- Barsbold, R.; Perle, A. (1980). "Segnosauria, a new suborder of carnivorous dinosaurs" (PDF). Acta Palaeontologica Polonica. 25 (2): 190−192.
- Barsbold, R. (1983). "O ptich'ikh chertakh v stroyenii khishchnykh dinozavrov" [Avian features in the morphology of predatory dinosaurs]. Transactions of the Joint Soviet Mongolian Paleontological Expedition (in Russian). 24: 96−103. Translated paper
- Paul, G. S. (1984). "The segnosaurian dinosaurs: relics of the prosauropod-ornithischian transition?". Journal of Vertebrate Paleontology. 4 (4): 507–515. doi:10.1080/02724634.1984.10012026. ISSN 0272-4634. JSTOR 4523011.
- Paul, G. S. (1988). Predatory Dinosaurs of the World. New York: Simon & Schuster. pp. 185−283. ISBN 9780671619466.
- Gauthier, J. (1986). "Saurischian monophyly and the origin of birds". Memoirs of the California Academy of Sciences. 8: 45.
- Sereno, P. C. (1989). "Prosauropod monophyly and basal sauropodomorph phylogeny". Abstract of Papers. Forty-Ninth Annual Meeting Society of Vertebrate Paleontology. Journal of Vertebrate Paleontology. 9 (supp. 3). p. 39A. ISSN 0272-4634. JSTOR 4523276.
- Norman, D. B. (1990). "Problematic Theropoda: Coelurosauria". In Weishampel, D. B.; Osmolska, H.; Dodson, P. (eds.). The Dinosauria (1st ed.). Berkeley: University of California Press. pp. 280−305. ISBN 9780520067271.
- Russell, D. A.; Dong, Z. (1993). "The affinities of a new theropod from the Alxa Desert, Inner Mongolia, People's Republic of China". Canadian Journal of Earth Sciences. 30 (10): 2107−2127. Bibcode:1993CaJES..30.2107R. doi:10.1139/e93-183.
- Clark, J. M.; Perle, A.; Norell, M. (1994). "The skull of Erlicosaurus andrewsi, a Late Cretaceous "Segnosaur" (Theropoda, Therizinosauridae) from Mongolia". American Museum Novitates. 3115: 1–39. hdl:2246/3712.
- Russell, D. A. (1997). "Therizinosauria". In Currie, P. J.; Padian, K. (eds.). Encyclopedia of Dinosaurs. San Diego: Academic Press. pp. 729−730. ISBN 978-0-12-226810-6.
- Xu, X.; Tang, Z.; Wang, X. A. (1999). "A therizinosauroid dinosaur with integumentary structures from China". Nature. 339 (6734): 350−354. Bibcode:1999Natur.399..350X. doi:10.1038/20670. S2CID 204993327.
- Hartman, S.; Mortimer, M.; Wahl, W. R.; Lomax, D. R.; Lippincott, J.; Lovelace, D. M. (2019). "A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight". PeerJ. 7: e7247. doi:10.7717/peerj.7247. PMC 6626525. PMID 31333906.
- Lautenschlager, S.; Emily, J. R.; Perle, A.; Lindsay, E. Z.; Lawrence, M. W. (2012). "The Endocranial Anatomy of Therizinosauria and Its Implications for Sensory and Cognitive Function". PLOS ONE. 7 (12): e52289. Bibcode:2012PLoSO...752289L. doi:10.1371/journal.pone.0052289. PMC 3526574. PMID 23284972.
- Kundrát, M.; Cruickshank, A. R. I.; Manning, T. W.; Nudds, J. (2007). "Embryos of therizinosauroid theropods from the Upper Cretaceous of China: diagnosis and analysis of ossification patterns". Acta Zoologica. 89 (3): 231–251. doi:10.1111/j.1463-6395.2007.00311.x.
- Kobayashi, Y.; Lee, Y.; Barsbold, R.; Zelenitsky, D.; Tanaka, K. (2013). "First record of a dinosaur nesting colony from Mongolia reveals nesting behavior of therizinosauroids". In Maxwell, E.; Miller-Camp, J.; Anemone, R. (eds.). 73rd Annual Meeting of Society of Vertebrate Paleontology. Los Angeles. p. 155.
- Tanaka, K.; Kobayashi, Y.; Zelenitsky, D. K.; Therrien, F.; Lee, Y.-N.; Barsbold, R.; Kubota, K.; Lee, H.-J.; Tsogtbaatar, C.; Idersaikhan, D. (2019). "Exceptional preservation of a Late Cretaceous dinosaur nesting site from Mongolia reveals colonial nesting behavior in a non-avian theropod". Geology. 47 (9): 843−847. Bibcode:2019Geo....47..843T. doi:10.1130/G46328.1.
- Nessov, L. A. (1995). Динозавры северной евразии: Новые данные о составе комплексов, экологии и палеобиогеографии [Dinosaurs of northern Eurasia: new data about assemblages, ecology, and paleobiogeography] (PDF) (in Russian). Saint Petersburg: Institute of Earth Crust, Saint Petersburg University. pp. 45−46. Translated paper
- Lautenschlager, S. (2014). "Morphological and functional diversity in therizinosaur claws and the implications for theropod claw evolution". Proceedings of the Royal Society B. 28 (1785): 20140497. doi:10.1098/rspb.2014.0497. PMC 4024305. PMID 24807260.
- Owocki, K.; Kremer, B.; Cotte, M.; Bocherens, H. (2020). "Diet preferences and climate inferred from oxygen and carbon isotopes of tooth enamel of Tarbosaurus bataar (Nemegt Formation, Upper Cretaceous, Mongolia)". Palaeogeography, Palaeoclimatology, Palaeoecology. 537: 109190. Bibcode:2020PPP...537j9190O. doi:10.1016/j.palaeo.2019.05.012.
- Eberth, D. A. (2018). "Stratigraphy and paleoenvironmental evolution of the dinosaur-rich Baruungoyot-Nemegt succession (Upper Cretaceous), Nemegt Basin, southern Mongolia". Palaeogeography, Palaeoclimatology, Palaeoecology. 494: 29−50. Bibcode:2018PPP...494...29E. doi:10.1016/j.palaeo.2017.11.018.
- Funston, G. F.; Mendonca, S. E.; Currie, P. J.; Barsbold, R.; Barsbold, R. (2018). "Oviraptorosaur anatomy, diversity and ecology in the Nemegt Basin". Palaeogeography, Palaeoclimatology, Palaeoecology. 494: 101−120. Bibcode:2018PPP...494..101F. doi:10.1016/j.palaeo.2017.10.023.
- Chinzorig, T.; Kobayashi, Y.; Tsogtbaatar, K.; Currie, P. J.; Takasaki, R.; Tanaka, T.; Iijima, M.; Barsbold, R. (2018). "Ornithomimosaurs from the Nemegt Formation of Mongolia: manus morphological variation and diversity". Palaeogeography, Palaeoclimatology, Palaeoecology. 494: 91–100. Bibcode:2018PPP...494...91C. doi:10.1016/j.palaeo.2017.10.031.
- Fanti, F.; Bell, P. R.; Currie, P. J.; Tsogtbataar, K. (2020). "The Nemegt Basin — One of the best field laboratories for interpreting Late Cretaceous terrestrial ecosystems". Palaeogeography, Palaeoclimatology, Palaeoecology. 494: 1–4. doi:10.1016/j.palaeo.2017.07.014.
- Holtz, T. R. (2014). "Mystery of the horrible hands solved". Nature. 515 (7526): 203−205. Bibcode:2014Natur.515..203H. doi:10.1038/nature13930. PMID 25337885.
External links
 Media related to Therizinosaurus at Wikimedia Commons
Media related to Therizinosaurus at Wikimedia Commons Data related to Therizinosaurus at Wikispecies
Data related to Therizinosaurus at Wikispecies- Therizinosaurus at the Encyclopædia Britannica
- 3D Therizinosaurus ungual at Sketchfab




.png.webp)



.jpg.webp)





