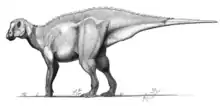
Ankylopollexia
Ankylopollexia is an extinct clade of ornithischian dinosaurs that lived from the Late Jurassic to the Late Cretaceous. It is a derived clade of iguanodontian ornithopods and contains the subgroup Styracosterna.[1] The name stems from the Greek word, “ankylos”, mistakenly taken to mean stiff, fused (in fact the adjective means bent or curved; used of fingers, it can mean hooked), and the Latin word, “pollex”, meaning thumb. Originally described in 1986 by Sereno, this most likely synapomorphic feature of a conical thumb spine defines the clade.[2]
| Ankylopollexia | |
|---|---|
 | |
| Mantellisaurus skeleton, Natural History Museum, London | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Order: | †Ornithischia |
| Suborder: | †Ornithopoda |
| Clade: | †Dryomorpha |
| Clade: | †Ankylopollexia Sereno, 1986 |
| Subgroups | |
| |
First appearing around 156 million years ago, in the Jurassic, Ankylopollexia became an extremely successful and widespread clade during the Cretaceous, and were found around the world. The group died out at the end of the Maastrichtian.[1] Even though they grew to be quite large, comparable to some carnivorous dinosaurs, they were universally herbivorous.[3]
Size

Ankylopollexians varied greatly in size over the course of their evolution.. Jurassic genus Camptosaurus was small, no more than 5 metres (16 ft) in length and half a tonne in weight.[4] The largest known ankylopollexian, dating to the late Campanian age (around 70 million years ago), belonged to the hadrosaurid family, and is named Shantungosaurus. It was around 14.7 metres (48 ft) to 16.6 metres (54 ft) in length and weighed, for the largest individuals, up to 16 tonnes (18 short tons).[5][6]

Primitive ankylopollexians tended to be smaller as compared to the larger, more derived hadrosaurs. There are, however, exceptions to this trend. A single track from a large ornithopod, likely a relative of Camptosaurus, was reported from the Lourinhã Formation, dating to the Jurassic in Portugal. The corresponding animal had an estimated hip height of around 2.8 metres (9.2 ft), much larger than the contemporary relative Draconyx.[7] The primitive styracosternan Iguanacolossus was named for its distinct robustness and large size, likely around 9 metres (30 ft) in length. Regarding hadrosaurs, one of the more basal members of Hadrosauroidea, the Chinese genus Bolong, is estimated to have been around 200 kilograms (440 lb).[8] Another exception of this trend is Tethyshadros, a more derived genus of Hadrosauroidea. Estimated to have weighed 350 kilograms (770 lb), Tethyshadros have been found only on certain islands in Italy. Its diminutive size is explained by insular dwarfism.[9]
Classification

About 157 million years ago, Ankylopollexia and Dryosauridae are believed to have split into separate evolutionary branches.[10] Originally named and described in 1986 by Paul Sereno, Ankylopollexia would receive a more formal definition in a later paper by Sereno in 2005.[2] In the 1986 paper, the groups Camptosauridae and Styracosterna were used to define the clade, but in the 2005 paper, a phylogenetic definition was given: the last common ancestor of the species Camptosaurus dispar and Parasaurolophus walkeri and all its descendants.
The cladogram below follows the phylogenetic analysis of Bertozzo et al. (2017).[11]
| Ankylopollexia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Palaeobiology
Brain
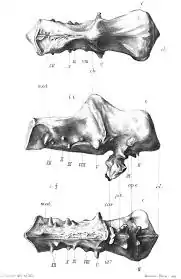
The neurobiology of ankylopollexians has been studied as far back as 1871, when a well preserved cranium (specimen NHMUK R2501[12]) discovered in September 1869 from the Wealden Group on the Isle of Wight and tentatively referred to the genus Iguanodon was described by John Hulke. He noted that due to the lesser correlation of the shape of the brain and wall of cranial cavity in reptiles, any deduction of the shape of the brain of the animal would be approximate.[13] The referral of this skull was reinforced in a later study, published in 1897. It was here inquired that the brain of the dinosaur may have been more closely associated to the cavity than that of modern reptiles, and so an endocast was created and studied.[14] This was not the first endocast of an ankylopolloxian brain, for in 1893, the skull of a Claosaurus annectens (today referred to the genus Edmontosaurus[15]) was used by Othniel Charles Marsh to create a cast of the brain cavity. Some basics remarks were made, including the small size of the organ, but interpreting minute features of the organ was noted to be difficult.[16] The 1897 paper noted the similarity of the two endocasts.[14]
Hadrosaurs have been noted as having the most complex brains among ankylopollexians, and indeed among ornithischian dinosaurs as a whole. The brains of a large variety of taxa have been studied. John Ostrom, would, in 1961, provide what was then the most extensive and detailed review and work on hadrosaur neuro-anatomy. This area of hadrosaur study was in its infancy at this point, and only the species known today as Edmontosaurus annectens, Edmontosaurus regalis, and Gryposaurus notabilis (at that time thought to be a synonym of its relative Kritosaurus) had specimens suitable at the time to be examined (Lambeosaurus was listed as having a briefly described braincase, but this was a mistake originating in Lull and Wright (1942)).[17][18] Ostrom supported the view that the brains of hadrosaurs and other dinosaurs would've likely only filled a portion of the cranial cavity, therefore hindering the ability to learn from endocasts, but noted they were still useful. He noted, similar to Marsh, noted the small predicted size of the organ, but also that it was significantly developed. A number of similarities to the brains of modern reptiles were noted.[18]

James Hopson investigated the encephalization quotients (EQs) of various dinosaurs in 1977 study. Three ornithopods for which brain endocasts had previously been produced – Camptosaurus, Iguanodon, and Anatosaurus (now known as Edmontosaurus annectens[15]) – were investigated. It was found that they had relatively high EQs compared to many other dinosaurs (ranging from 0.8 to 1.5), comparable to that of carnosaurian theropods and of modern crocodilians, but far lower than that of coelurosaurian theropods. The latter two genera, which lived later than Camptosaurus, had somewhat higher EQs than the Jurassic taxon, which, being at the lower end, was more comparable to the ceratopsian genus Protoceratops. Reasonings suggested for their comparably high intelligence were the need for acute senses in the lack of defensive weapons, and more complex intraspecific behaviours as indicated by their acoustic and visual display structures.[19]
In a first for any terrestrial fossil vertebrate, Brasier et al. (2017) reported mineralized soft tissues from the brain of an iguanodontian dinosaur, from the Valanginian age (around 133 million years ago) Upper Tunbridge Wells Formation at Bexhill, Sussex. Fragmentary ornithopod remains were associated with the fossil, and though assigning the specimen to any one taxon with certainty wasn't possible, Barilium or Hypselospinus were put forward as likely candidates. The specimen compared well to endocasts of similar taxa, such as one from a Mantellisaurus on display at the Oxford University Museum of Natural History. Detailed observations were made with the use of a scanning electron microscope. Only some parts of the brain were preserved; the cerebellar and celebral expansions were best preserved, whereas the olfactory lobes and medulla oblongata were missing or nearly so. The neural tissues seemed to be very tightly packed, indicating an EC closer to five (with hadrosaurs having even higher ECs), nearly matching that of the most intelligent non-avian theropods. Though it was noted this was in-line with their complex behaviour, as had been noted by Hopson, it was cautioned the dense packing may have been an artifact of preservation, and the original lower estimates were considered more accurate. Some of the complex behaviours ascribed can be seen to some extent in modern crocodilians, who fall near the original numbers.[12]

The advent of CT scanning for use in palaeontology has allowed for more widespread application of this without the need for specimen destruction. Modern research using these methods has focused largely on hadrosaurs. In a 2009 study by palaeontologist David C. Evans and colleagues, the brains of lambeosaurine hadrosaur genera Hypacrosaurus (adult specimen ROM 702), Corythosaurus (juvenile specimen ROM 759 and subadult specimen CMN 34825), and Lambeosaurus (juvenile specimen ROM 758) were scanned and compared to each other (on a phylogenetic and ontogenetic level), related taxa, and previous predictions, the first such large-scale look into the neurology of the subfamily. Contra the early works, Evans' studies indicate that only some regions of the hadrosaur brain (the dorsal portion and much of the hindbrain) were loosely correlated to the brain wall, like modern reptiles, with the ventral and lateral regions correlating fairly closely. Also unlike modern reptiles, the brains of the juveniles did not seem to correlate any closer to the brain wall than those of adults. It was cautioned, however, that very young individuals were not included in the study.[17]
As with previous studies, EQ values were investigated, although a wider number range was given to account for uncertainty in brain and body mass. The range for the adult Hypacrosaurus was 2.3 to 3.7; the lowest end of this range was still higher than modern reptiles and most non-maniraptoran dinosaurs (nearly all having EQs below two), but fell well short of maniraptorans themselves, which had quotients higher than four. The size of the cerebral hemispheres was, for the first time, remarked upon. It was found to taking up around 43% of endocranial volume (not considering olfactory bulbs) in ROM 702. This is comparable to their size in saurolophine hadrosaurs, but far larger than in any ornithischians outside of Hadrosauriformes, and all large saurischian dinosaurs; maniraptors Conchoraptor and Archaeopteryx, an early bird, had very similar proportions. This lends further support to the idea of complex behaviours and relatively high intelligence, for non-avian dinosaurs, in hadrosaurids.[17]

Amurosaurus, a close relative of the taxa from the 2009 study, was the subject of a 2013 paper once again looking into a cranial endocast. A nearly identical EQ range of 2.3 to 3.8 was found, and it was again noted this was higher than that of living reptiles, sauropods and other ornithischians, but different EQ estimates for theropods were cited, placing the hadrosaur numbers significantly below even more basal theropods like Ceratosaurus (with an EQ range of 3.31 to 5.07) and Allosaurus (with a range of 2.4 to 5.24, compared to only 1.6 in the 2009 study[17][20]); more bird-like coelurosaurians theropods such as Troodon had stated EQs higher than seven. Additionally, the relative cerebral volume was only 30% in Amurosaurus, significantly lower than in Hypacrosaurus, closer to that of theropods like Tyrannosaurus (with 33%), though still distinctly larger than previously estimated numbers for more primitive iguanodonts like Lurdusaurus and Iguanodon (both at 19%). This demonstrated a previously unrecognized level of variation in neuro-anatomy within Hadrosauridae.[20]
Palaeobiogeography

Ankylopollexians would in the Cretaceous become one of the most successful groups on the planet, being both widespread and numerous in nature.[21] Around this time, ankylopollexians spread to Asia and Europe. An early example is the Chinese genus Bayannurosaurus, from the Berriasian.[22] The oldest genus, found in Wyoming, is Camptosaurus dispar, which dates to around the Callovian-Oxfordian, about 156-157 million years ago.[23]
References
- McDonald, A. T. (2012). Farke, Andrew A (ed.). "Phylogeny of Basal Iguanodonts (Dinosauria: Ornithischia): An Update". PLOS ONE. 7 (5): e36745. Bibcode:2012PLoSO...736745M. doi:10.1371/journal.pone.0036745. PMC 3358318. PMID 22629328.
- Sereno, P.C. (1986). "Phylogeny of the bird-hipped dinosaurs (order Ornithischia)". National Geographic Research 2 (2): 234–56
- Foster, J. (2007). Camptosaurus dispar. Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. p. 219-221
- Paul, G.S., 2010, The Princeton Field Guide to Dinosaurs, Princeton University Press p. 284
- Glut, Donald F. (1997). "Shantungosaurus". Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. pp. 816–817. ISBN 0-89950-917-7.
- Zhao, X.; Li, D.; Han, G.; Hao, H.; Liu, F.; Li, L.; Fang, X. (2007). "Zhuchengosaurus maximus from Shandong Province". Acta Geoscientia Sinica 28 (2): 111–122. doi:10.1007/s10114-005-0808-x.
- Mateus, Octávio; Milàn, Jesper (2008). "Ichnological evidence for giant ornithopod dinosaurs in the Upper Jurassic Lourinhã Formation, Portugal". Oryctos. 8: 47–52.
- Wu Wen-hao, Pascal Godefroit, Hu Dong-yu (2010). "Bolong yixianensis gen. et sp. nov.: A new Iguanodontoid dinosaur from the Yixian Formation of Western Liaoning, China". Geology and Resources 19 (2): 127–133.
- Dalla Vecchia, F. M. (2009). "Tethyshadros insularis, a new hadrosauroid dinosaur (Ornithischia) from the Upper Cretaceous of Italy". Journal of Vertebrate Paleontology 29 (4): 1100–1116.
- Norman, David B.; Weishampel, David B. (1990). "Iguanodontidae and related ornithopods". In Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). The Dinosauria. Berkeley: University of California Press. pp. 510–533. ISBN 0-520-06727-4.
- Bertozzo, Filippo; Dalla Vecchia, Fabio Marco; Fabbri, Matteo (2017). "The Venice specimen of Ouranosaurus nigeriensis (Dinosauria, Ornithopoda)". PeerJ. 5 (e3403): e3403. doi:10.7717/peerj.3403. PMC 5480399. PMID 28649466.
- Brasier, Martin D.; et al. (2017). "Remarkable preservation of brain tissues in an Early Cretaceous iguanodontian dinosaur". Geological Society, London, Special Publications. 448 (1): 383–398. Bibcode:2017GSLSP.448..383B. doi:10.1144/SP448.3.
- Hulke J.W. (1871). "Note on a large reptilian skull from Brooke, Isle of Wight, probably Dinosaurian, referable to the genus Iguanodon". Quart J. Geol Soc. 27 (1–2): 199–206. doi:10.1144/GSL.JGS.1871.027.01-02.27. S2CID 130674982.
- Andrews, Chas. W. (1897). "Note on a cast of the brain-cavity of Iguanodon". Journal of Natural History. 19 (114): 585–591. doi:10.1080/00222939708680580.
- Creisler, Benjamin S. (2007). "Deciphering duckbills: a history in nomenclature". In Carpenter Kenneth (ed.). Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington and Indianapolis: Indiana University Press. pp. 185–210. ISBN 978-0-253-34817-3.
- Marsh, Othniel Charles (1893). "The skull and brain of Claosaurus". American Journal of Science. 265 (265): 83–86. Bibcode:1893AmJS...45...83M. doi:10.2475/ajs.s3-45.265.83. S2CID 131740074.
- Evans, David C.; et al. (2009). "Endocranial Anatomy of Lambeosaurine Hadrosaurids (Dinosauria: Ornithischia): A Sensorineural Perspective on Cranial Crest Function". The Anatomical Record. 292 (9): 1315–1337. doi:10.1002/ar.20984. PMID 19711466. S2CID 15177074.
- Ostrom, John H. (1961). "Cranial morphology of the hadrosaurian dinosaurs of North America". Bulletin of the AMNH. 122. hdl:2246/1260.
- Hopson, James A. (1977). "Relative brain size and behavior in archosaurian reptiles". Annual Review of Ecology and Systematics. 8 (1): 429–448. doi:10.1146/annurev.es.08.110177.002241.
- Lauters, Pascaline; et al. (2013). "Cranial Endocast of the Lambeosaurine Hadrosaurid Amurosaurus riabinini from the Amur Region, Russia". PLOS ONE. 8 (11): e78899. Bibcode:2013PLoSO...878899L. doi:10.1371/journal.pone.0078899. PMC 3827337. PMID 24236064.
- MacDonald, Andrew; et al. (2010). "New Basal Iguanodonts from the Cedar Mountain Formation of Utah and the Evolution of Thumb-Spiked Dinosaurs". PLOS ONE. 5 (11): e14075. Bibcode:2010PLoSO...514075M. doi:10.1371/journal.pone.0014075. PMC 2989904. PMID 21124919.
- Xu, Xing; et al. (2018). "A large-sized basal ankylopollexian from East Asia, shedding light on early biogeographic history of Iguanodontia". Science Bulletin. 63 (9): 556–563. doi:10.1016/j.scib.2018.03.016.
- Carpenter, K. and Wilson, Y. (2008). "A new species of Camptosaurus (Ornithopoda: Dinosauria) from the Morrison Formation (Upper Jurassic) of Dinosaur National Monument, Utah, and a biomechanical analysis of its forelimb". Annals of the Carnegie Museum 76: 227–263. doi:10.2992/0097-4463(2008)76[227:ansoco]2.0.co;2.
External links
 Data related to Iguanodontia at Wikispecies
Data related to Iguanodontia at Wikispecies
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||



.jpg.webp)