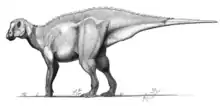
Edmontosaurus
Edmontosaurus (/ɛdˌmɒntəˈsɔːrəs/ ed-MON-tə-SAWR-əs) (meaning "lizard from Edmonton") is a genus of hadrosaurid (duck-billed) dinosaur. It contains two known species: Edmontosaurus regalis and Edmontosaurus annectens. Fossils of E. regalis have been found in rocks of western North America that date from the late Campanian stage of the Cretaceous Period 73 million years ago, while those of E. annectens were found in the same geographic region but in rocks dated to the end of the Maastrichtian stage of the Cretaceous, 66 million years ago. Edmontosaurus was one of the last non-avian dinosaurs, and lived alongside dinosaurs like Triceratops, Tyrannosaurus, Albertosaurus and Pachycephalosaurus shortly before the Cretaceous–Paleogene extinction event.
| Edmontosaurus | |
|---|---|
 | |
| Mounted cast of a fossil E. annectens skeleton, Rocky Mountain Dinosaur Resource Center | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Order: | †Ornithischia |
| Suborder: | †Ornithopoda |
| Family: | †Hadrosauridae |
| Subfamily: | †Saurolophinae |
| Tribe: | †Edmontosaurini |
| Genus: | †Edmontosaurus Lambe, 1917 |
| Type species | |
| †Edmontosaurus regalis Lambe, 1917 | |
| Other species | |
| Synonyms | |
| |
Edmontosaurus included some of the largest hadrosaurid species, measuring up to 12 metres (39 ft) long and weighing around 4.0 metric tons (4.4 short tons). Evidence does exist in the form of two fossilized specimens housed at the Museum of the Rockies for an even greater maximum size of 15 m (49 ft) and weighing 9.07 metric tons (10.00 short tons) for Edmontosaurus annectens.[2] Several well-preserved specimens are known that include not only bones, but in some cases extensive skin impressions and possible gut contents. It is classified as a genus of saurolophine (or hadrosaurine) hadrosaurid, a member of the group of hadrosaurids which lacked large, hollow crests, instead having smaller solid crests or fleshy combs.[3]
The first fossils named Edmontosaurus were discovered in southern Alberta (named after Edmonton, the capital city), in the Horseshoe Canyon Formation (formerly called the lower Edmonton Formation). The type species, E. regalis, was named by Lawrence Lambe in 1917, although several other species that are now classified in Edmontosaurus were named earlier. The best known of these is E. annectens, named by Othniel Charles Marsh in 1892; originally as a species of Claosaurus, known for many years as a species of Trachodon, and later as Anatosaurus annectens. Anatosaurus, Anatotitan and Ugrunaaluk are now generally regarded as synonyms of Edmontosaurus.
Edmontosaurus was widely distributed across western North America. The distribution of Edmontosaurus fossils suggests that it preferred coasts and coastal plains. It was a herbivore that could move on both two legs and four. Because it is known from several bone beds, Edmontosaurus is thought to have lived in groups, and may have been migratory as well. The wealth of fossils has allowed researchers to study its paleobiology in detail, including its brain, how it may have fed, and its injuries and pathologies, such as evidence for tyrannosaur attacks on a few edmontosaur specimens.
Discovery and history
Claosaurus annectens
Edmontosaurus has had a long and complicated history in paleontology, having spent decades with various species classified in other genera. Its taxonomic history intertwines at various points with the genera Agathaumas, Anatosaurus, Anatotitan, Claosaurus, Hadrosaurus, Thespesius, and Trachodon,[4][5] and references predating the 1980s typically use Anatosaurus, Claosaurus, Thespesius, or Trachodon for edmontosaur fossils (excluding those assigned to E. regalis), depending on author and date. Although Edmontosaurus was only named in 1917, its oldest well-supported species (E. annectens) was named in 1892 as a species of Claosaurus.
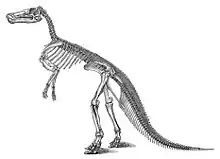
The first well-supported species of Edmontosaurus was named in 1892 as Claosaurus annectens by Othniel Charles Marsh. This species is based on USNM 2414, a partial skull-roof and skeleton, with a second skull and skeleton, YPM 2182, designated the paratype. Both were collected in 1891 by John Bell Hatcher from the late Maastrichtian-age Upper Cretaceous Lance Formation of Niobrara County (then part of Converse County), Wyoming.[6] This species has some historical footnotes attached: it is among the first dinosaurs to receive a skeletal restoration, and is the first hadrosaurid so restored;[5][7] and YPM 2182 and UNSM 2414 are, respectively, the first and second essentially complete mounted dinosaur skeletons in the United States.[8] YPM 2182 was put on display in 1901,[5] and USNM 2414 in 1904.[8]

Because of the incomplete understanding of hadrosaurids at the time, following Marsh's death in 1897 Claosaurus annectens was variously classified as a species of Claosaurus, Thespesius or Trachodon. Opinions varied greatly; textbooks and encyclopedias drew a distinction between the "Iguanodon-like" Claosaurus annectens and the "duck-billed" Hadrosaurus (based on remains now known as adult Edmontosaurus annectens), while Hatcher explicitly identified C. annectens as synonymous with the hadrosaurid represented by those same duck-billed skulls.[5] Hatcher's revision, published in 1902, was sweeping: he considered almost all hadrosaurid genera then known as synonyms of Trachodon. This included Cionodon, Diclonius, Hadrosaurus, Ornithotarsus, Pteropelyx, and Thespesius, as well as Claorhynchus and Polyonax, fragmentary genera now thought to be horned dinosaurs.[9] Hatcher's work led to a brief consensus, until after 1910 new material from Canada and Montana showed a greater diversity of hadrosaurids than previously suspected.[5] Charles W. Gilmore in 1915 reassessed hadrosaurids and recommended that Thespesius be reintroduced for hadrosaurids from the Lance Formation and rock units of equivalent age, and that Trachodon, based on inadequate material, should be restricted to a hadrosaurid from the older Judith River Formation and its equivalents. In regards to Claosaurus annectens, he recommended that it be considered the same as Thespesius occidentalis.[10] His reinstatement of Thespesius for Lance-age hadrosaurids would have other consequences for the taxonomy of Edmontosaurus in the following decades.
During this time frame (1902–1915), two additional important specimens of C. annectens were recovered. The first, the "mummy" specimen AMNH 5060, was discovered in 1908 by Charles Hazelius Sternberg and his sons in Lance Formation rocks near Lusk, Wyoming. Sternberg was working for the British Museum of Natural History, but Henry Fairfield Osborn of the American Museum of Natural History was able to purchase the specimen for $2,000.[11] The Sternbergs recovered a second similar specimen from the same area in 1910,[12] not as well preserved but also found with skin impressions. They sold this specimen (SM 4036) to the Senckenberg Museum in Germany.[11]
As a side note, Trachodon selwyni, described by Lawrence Lambe in 1902 for a lower jaw from what is now known as the Dinosaur Park Formation of Alberta,[13] was erroneously described by Glut (1997) as having been assigned to Edmontosaurus regalis by Lull and Wright.[14] It was not, instead being designated "of very doubtful validity."[15] More recent reviews of hadrosaurids have concurred.[4][16]
Canadian discoveries
Edmontosaurus itself was coined in 1917 by Lawrence Lambe for two partial skeletons found in the Horseshoe Canyon Formation (formerly the lower Edmonton Formation) along the Red Deer River of southern Alberta, Canada.[17] These rocks are older than the rocks in which Claosaurus annectens was found.[18] The Edmonton Formation lends Edmontosaurus its name.[5] The type species, E. regalis ("regal", or, more loosely, "king-sized"),[5] is based on NMC 2288, consisting of a skull, articulated vertebrae up to the sixth tail vertebra, ribs, partial hips, an upper arm bone, and most of a hind limb. It was discovered in 1912 by Levi Sternberg. The second specimen, paratype NMC 2289, consists of a skull and skeleton lacking the beak, most of the tail, and part of the feet. It was discovered in 1916 by George F. Sternberg. Lambe found that his new dinosaur compared best to Diclonius mirabilis (specimens now assigned to Edmontosaurus annectens), and drew attention to the size and robustness of Edmontosaurus.[17] Initially, Lambe only described the skulls of the two skeletons, but returned to the genus in 1920 to describe the skeleton of NMC 2289.[19] The postcrania of the type specimen remains undescribed, still in its plaster jackets.[14]

Two more species that would come to be included with Edmontosaurus were named from Canadian remains in the 1920s, but both would initially be assigned to Thespesius. Gilmore named the first, Thespesius edmontoni, in 1924. T. edmontoni also came from the Horseshoe Canyon Formation. It was based on NMC 8399, another nearly complete skeleton lacking most of the tail. NMC 8399 was discovered on the Red Deer River in 1912 by a Sternberg party.[20] Its forelimbs, ossified tendons, and skin impressions were briefly described in 1913 and 1914 by Lambe, who at first thought it was an example of a species he had named Trachodon marginatus,[21] but then changed his mind.[22] The specimen became the first dinosaur skeleton to be mounted for exhibition in a Canadian museum. Gilmore found that his new species compared closely to what he called Thespesius annectens, but left the two apart because of details of the arms and hands. He also noted that his species had more vertebrae than Marsh's in the back and neck, but proposed that Marsh was mistaken in assuming that the annectens specimens were complete in those regions.[20]
In 1926, Charles Mortram Sternberg named Thespesius saskatchewanensis for NMC 8509, a skull and partial skeleton from the Wood Mountain plateau of southern Saskatchewan. He had collected this specimen in 1921, from rocks that were assigned to the Lance Formation,[23] now the Frenchman Formation.[4] NMC 8509 included an almost complete skull, numerous vertebrae, partial shoulder and hip girdles, and partial hind limbs, representing the first substantial dinosaur specimen recovered from Saskatchewan. Sternberg opted to assign it to Thespesius because that was the only hadrosaurid genus known from the Lance Formation at the time.[23] At the time, T. saskatchewanensis was unusual because of its small size, estimated at 7 to 7.3 metres (23 to 24 ft) in length.[24]
Anatosaurus to the present

In 1942, Lull and Wright attempted to resolve the complicated taxonomy of crestless hadrosaurids by naming a new genus, Anatosaurus, to take in several species that did not fit well under their previous genera. Anatosaurus, meaning "duck lizard", because of its wide, duck-like beak (Latin anas = duck + Greek sauros = lizard), had as its type species Marsh's old Claosaurus annectens. Also assigned to this genus were Thespesius edmontoni, T. saskatchewanensis, a large lower jaw that Marsh had named Trachodon longiceps in 1890, and a new species, Anatosaurus copei, for two skeletons on display at the American Museum of Natural History that had long been known as Diclonius mirabilis (or variations thereof). Thus, the various species became Anatosaurus annectens, A. copei, A. edmontoni, A. longiceps, and A. saskatchewanensis.[24] Anatosaurus would come to be called the "classic duck-billed dinosaur."[25]

This state of affairs persisted for several decades, until Michael K. Brett-Surman reexamined the pertinent material for his graduate studies in the 1970s and 1980s. He concluded that the type species of Anatosaurus, A. annectens, was actually a species of Edmontosaurus and that A. copei was different enough to warrant its own genus.[26][27][28] Although theses and dissertations are not regarded as official publications by the International Commission on Zoological Nomenclature, which regulates the naming of organisms, his conclusions were known to other paleontologists, and were adopted by several popular works of the time.[29][30] Brett-Surman and Ralph Chapman designated a new genus for A. copei (Anatotitan) in 1990.[31] Of the remaining species, A. saskatchewanensis and A. edmontoni were assigned to Edmontosaurus as well,[16] and A. longiceps went to Anatotitan, as either a second species[32] or as a synonym of A. copei.[16] Because the type species of Anatosaurus (A. annectens) was sunk into Edmontosaurus, the name Anatosaurus is abandoned as a junior synonym of Edmontosaurus.
The conception of Edmontosaurus that emerged included three valid species: the type E. regalis, E. annectens (including Anatosaurus edmontoni, amended to edmontonensis), and E. saskatchewanensis.[16] The debate about the proper taxonomy of the A. copei specimens continues to the present: returning to Hatcher's argument of 1902, Jack Horner, David B. Weishampel, and Catherine Forster regarded Anatotitan copei as representing specimens of Edmontosaurus annectens with crushed skulls.[4] In 2007 another "mummy" was announced; nicknamed "Dakota", it was discovered in 1999 by Tyler Lyson, and came from the Hell Creek Formation of North Dakota.[33][34]
In a 2011 study by Nicolás Campione and David Evans, the authors conducted the first ever morphometric analysis to compare the various specimens assigned to Edmontosaurus. They concluded that only two species are valid: E. regalis, from the late Campanian, and E. annectens, from the late Maastrichtian. Their study provided further evidence that Anatotitan copei is a synonym of E. annectens; specifically, that the long, low skull of A. copei is the result of ontogenetic change and represents mature E. annectens individuals.[18]
Species and distribution
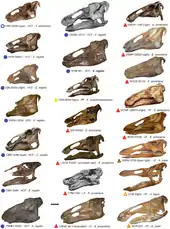
Edmontosaurus is currently regarded as having two valid species: type species E. regalis, and E. annectens.[4][18] E. regalis is known only from the Horseshoe Canyon Formation of Alberta, dating from the late Campanian stage of the late Cretaceous period. At least a dozen individuals are known,[18] including seven skulls with associated postcrania, and five to seven other skulls.[4][16] The species formerly known as Thespesius edmontoni or Anatosaurus edmontoni represents immature individuals.[18][35][36]
E. annectens is known from the Frenchman Formation of Saskatchewan, the Hell Creek Formation of Montana, and the Lance Formation of South Dakota and Wyoming. It is limited to late Maastrichtian rocks, and is represented by at least twenty skulls, some with postcranial remains.[18] One author, Kraig Derstler, has described E. annectens as "perhaps the most perfectly-known dinosaur to date [1994]."[37] Anatosaurus copei and E. saskatchewanensis are now thought to be growth stages of E. annectens: A. copei as adults, and E. saskatchewanensis as juveniles.[18] Trachodon longiceps may be a synonym of E. annectens as well.[4] Anatosaurus edmontoni was mistakenly listed as a synonym of E. annectens in both reviews of the Dinosauria,[4][16] but this does not appear to be the case.[18][36]

E. annectens differed from E. regalis by having a longer, lower, less robust skull.[14][18] Although Brett-Surman regarded E. regalis and E. annectens as potentially representing males and females of the same species,[26] all E. regalis specimens come from older formations than E. annectens specimens.[36] Edmontosaurine specimens from the Prince Creek Formation of Alaska formerly assigned to Edmontosaurus sp. have been given their own genus and species name, Ugrunaaluk kuukpikensis.[38] However, the identification of Ugrunaaluk as a separate genus was questioned by a 2017 study from Hai Xing and colleagues, who regarded it as a nomen dubium indistinguishable from other Edmontosaurus.[1] In 2020, Ryuji Takasaki and colleagues agreed that the Prince Creek remains should be classified as Edmontosaurus, though species designation is unclear as the specimens are juveniles.[39] Edmontosaurus was also reported from the Javelina Formation of Big Bend National Park, western Texas based on TMM 41442-1,[40] but was later referred to Kritosaurus cf. navajovius by Wagner (2001), before being assigned to Kritosaurus sp. by Lehman et al. (2016).[41][42][43]
Description
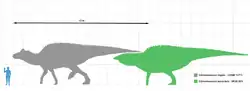
Edmontosaurus has been described in detail from numerous specimens.[19][20][23][44] Like other hadrosaurids, it was a bulky animal with a long, laterally flattened tail and a head with an expanded, duck-like beak. The fore legs were not as heavily built as the hind legs, but were long enough to be used in standing or movement. Edmontosaurus was among the largest hadrosaurids: depending on the species, a fully grown adult could have been 9 metres (30 ft) long, and some of the larger specimens reached the range of 12 metres (39 ft)[14] to 13 metres (43 ft) long.[45] Its weight was on the order of 4.0 metric tons (4.4 short tons).[4] Traditionally, E. regalis has been regarded as the largest species, though this was challenged by the hypothesis that the larger hadrosaurid Anatotitan copei is a synonym of Edmontosaurus annectens, as put forward by Jack Horner and colleagues in 2004,[4] and supported in studies by Campione and Evens in 2009 and 2011.[18] The type specimen of E. regalis, NMC 2288, is estimated as 9 to 12 metres (30 to 39 ft) long.[46] E. annectens is often seen as smaller. Two mounted skeletons, USNM 2414 and YPM 2182, measure 8.00 metres (26.25 ft) long and 8.92 metres (29.3 ft) long, respectively.[46][8] However, these are probably subadult individuals,[18] and there is at least one report of a much larger potential E. annectens specimen, almost 12 metres (39 ft) long.[47] Two specimens still under study in the collection of the Museum of the Rockies - a 7.6 m (25 ft) tail labelled as MOR 1142 and another labelled as MOR 1609 - indicate that Edmontosaurus annectens could have grown to larger sizes, possibly rivaling Shantungosaurus in size. The specimens of these individuals indicate a length of up to 15 m (49 ft), according to paleontologists.[2] Such large individuals of Edmontosaurus would likely have been very rare due to such factors as environmental stress, disease, and predation.
Skull

The skull of a fully grown Edmontosaurus could be over a metre long. One skull of E. annectens (formerly Anatotitan) measures 3.87 feet (1.18 m) long.[48] The skull was roughly triangular in profile,[19] with no bony cranial crest.[24] Viewed from above, the front and rear of the skull were expanded, with the broad front forming a duck-bill or spoon-bill shape. The beak was toothless, and both the upper and lower beaks were extended by keratinous material.[4] Substantial remains of the keratinous upper beak are known from the "mummy" kept at the Senckenberg Museum.[14] In this specimen, the preserved nonbony part of the beak extended for at least 8 centimetres (3.1 in) beyond the bone, projecting down vertically.[49] The nasal openings of Edmontosaurus were elongate and housed in deep depressions surrounded by distinct bony rims above, behind, and below.[35] In at least one case (the Senckenberg specimen), rarely preserved sclerotic rings were preserved in the eye sockets.[50] Another rarely seen bone, the stapes (the reptilian ear bone), has also been seen in a specimen of Edmontosaurus.[4]
Teeth were present only in the maxillae (upper cheeks) and dentaries (main bone of the lower jaw). The teeth were continually replaced, taking about half a year to form.[51] They were composed of six types of tissues, rivaling the complexity of mammal teeth.[52] They grew in columns, with an observed maximum of six in each, and the number of columns varied based on the animal's size.[44] Known column counts for the two species are: 51 to 53 columns per maxilla and 48 to 49 per dentary (teeth of the upper jaw being slightly narrower than those in the lower jaw) for E. regalis; and 52 columns per maxilla and 44 per dentary for E. annectens (an E. saskatchewanensis specimen).[24]
Postcranial skeleton

The number of vertebrae differs between specimens. E. regalis had thirteen neck vertebrae, eighteen back vertebrae, nine hip vertebrae, and an unknown number of tail vertebrae.[24] A specimen once identified as belonging to Anatosaurus edmontoni (now considered to be the same as E. regalis) is reported as having an additional back vertebra and 85 tail vertebrae, with an undisclosed amount of restoration.[24] Other hadrosaurids are only reported as having 50 to 70 tail vertebrae,[4] so this appears to have been an overestimate. The anterior back was curved toward the ground, with the neck flexed upward and the rest of the back and tail held horizontally.[4] Most of the back and tail were lined by ossified tendons arranged in a latticework along the neural spines of the vertebrae. This condition has been described as making the back and at least part of the tail "ramrod" straight.[53][54] The ossified tendons are interpreted as having strengthened the vertebral column against gravitational stress, incurred through being a large animal with a horizontal vertebral column otherwise supported mostly by the hind legs and hips.[53]
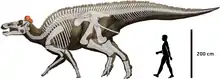
The shoulder blades were long flat blade-like bones, held roughly parallel to the vertebral column. The hips were composed of three elements each: an elongate ilium above the articulation with the leg, an ischium below and behind with a long thin rod, and a pubis in front that flared into a plate-like structure. The structure of the hip hindered the animal from standing with its back erect, because in such a position the thigh bone would have pushed against the joint of the ilium and pubis, instead of pushing only against the solid ilium. The nine fused hip vertebrae provided support for the hip.[44]
The fore legs were shorter and less heavily built than the hind legs. The upper arm had a large deltopectoral crest for muscle attachment, while the ulna and radius were slim. The upper arm and forearm were similar in length. The wrist was simple, with only two small bones. Each hand had four fingers, with no thumb (first finger). The index (second), third, and fourth fingers were approximately the same length and were united in life within a fleshy covering. Although the second and third finger had hoof-like unguals, these bones were also within the skin and not apparent from the outside. The little finger diverged from the other three and was much shorter. The thigh bone was robust and straight, with a prominent flange about halfway down the posterior side.[44] This ridge was for the attachment of powerful muscles attached to the hips and tail that pulled the thighs (and thus the hind legs) backward and helped maintain the use of the tail as a balancing organ.[55] Each foot had three toes, with no big toe or little toe. The toes had hoof-like tips.[44]
Soft tissue

Multiple specimens of Edmontosaurus annectens have been found with preserved skin impressions. Several have been well-publicized, such as the "Trachodon mummy" of the early 20th century,[56][57] and the specimen nicknamed "Dakota",[33][34][58] the latter apparently including remnant organic compounds from the skin.[58] Because of these finds, the scalation of Edmontosaurus annectens is known for most areas of the body. Skin impressions are less well known for E. regalis, but some well-preserved examples have been studied, including one which preserves a soft tissue crest or wattle on the head. It is unknown whether such a crest was present on E. annectens, and whether it was an indicator of sexual dimorphism.[3]
A preserved rhamphotheca present in the E. annectens specimen LACM 23502, housed in the Los Angeles County Museum, indicates the beak of Edmontosaurus was more hook-shaped and extensive than many illustrations in scientific and public media have previously depicted. Whether or not the specimen in question preserved the true rhamphotheca or just a cast of the inner structure attached to the bone is not known at present.[59][60][61]
Classification
Edmontosaurus was a hadrosaurid (a duck-billed dinosaur), a member of a family of dinosaurs which to date are known only from the Late Cretaceous. It is classified within the Saurolophinae (alternately Hadrosaurinae), a clade of hadrosaurids which lacked hollow crests. Other members of the group include Brachylophosaurus, Gryposaurus, Lophorhothon, Maiasaura, Naashoibitosaurus, Prosaurolophus, and Saurolophus.[4] It was either closely related to[16] or includes the species Anatosaurus annectens (alternately Edmontosaurus annectens),[4] a large hadrosaurid from various latest Cretaceous formations of western North America. The giant Chinese hadrosaurine Shantungosaurus giganteus is also anatomically similar to Edmontosaurus; M. K. Brett-Surman found the two to differ only in details related to the greater size of Shantungosaurus, based on what had been described of the latter genus.[26]
While the status of Edmontosaurus as a saurolophine has not been challenged, its exact placement within the clade is uncertain. Early phylogenies, such as that presented in R. S. Lull and Nelda Wright's influential 1942 monograph, had Edmontosaurus and various species of Anatosaurus (most of which would be later considered as additional species or specimens of Edmontosaurus) as one lineage among several lineages of "flat-headed" hadrosaurs.[62] One of the first analyses using cladistic methods found it to be linked with Anatosaurus (=Anatotitan) and Shantungosaurus in an informal "edmontosaur" clade, which was paired with the spike-crested "saurolophs" and more distantly related to the "brachylophosaurs" and arch-snouted "gryposaurs".[16] A 2007 study by Terry Gates and Scott Sampson found broadly similar results, in that Edmontosaurus remained close to Saurolophus and Prosaurolophus and distant from Gryposaurus, Brachylophosaurus, and Maiasaura.[63] However, the most recent review of Hadrosauridae, by Jack Horner and colleagues (2004), came to a noticeably different result: Edmontosaurus was nested between Gryposaurus and the "brachylophosaurs", and distant from Saurolophus.[4]
Left cladogram per Horner et al. (2004),[4] right cladogram per Gates and Sampson (2007).[63]
|
|
Paleobiology

Growth
In a 2011 study, Campione and Evans recorded data from all known "edmontosaur" skulls from the Campanian and Maastrichtian and used it to plot a morphometric graph, comparing variable features of the skull with skull size. Their results showed that within both recognized Edmontosaurus species, many features previously used to classify additional species or genera were directly correlated with skull size. Campione and Evans interpreted these results as strongly suggesting that the shape of Edmontosaurus skulls changed dramatically as they grew. This has led to several apparent mistakes in classification in the past. The Campanian species Thespesius edmontoni, previously considered a synonym of E. annectens due to its small size and skull shape, is more likely a subadult specimen of the contemporary E. regalis. Similarly, the three previously recognized Maastrichtian edmontosaur species likely represent growth stages of a single species, with E. saskatchewanensis representing juveniles, E. annectens subadults, and Anatotitan copei fully mature adults. The skulls became longer and flatter as the animals grew.[18]
Brain and nervous system

The brain of Edmontosaurus has been described in several papers and abstracts through the use of endocasts of the cavity where the brain had been. E. annectens[64][65] and E. regalis,[19] as well as specimens not identified to species,[66][67][68] have been studied in this way. The brain was not particularly large for an animal the size of Edmontosaurus. The space holding it was only about a quarter of the length of the skull,[19] and various endocasts have been measured as displacing 374 millilitres (13 US fl oz)[68] to 450 millilitres (15 US fl oz),[67] which does not take into account that the brain may have occupied as little as 50% of the space of the endocast, the rest of the space being taken up by the dura mater surrounding the brain.[67][68] For example, the brain of the specimen with the 374 millilitre endocast is estimated to have had a volume of 268 millilitres (9 US fl oz).[68] The brain was an elongate structure,[67] and as with other non-mammals, there would have been no neocortex.[68] Like Stegosaurus, the neural canal was expanded in the hips, but not to the same degree: the endosacral space of Stegosaurus had 20 times the volume of its endocranial cast, whereas the endosacral space of Edmontosaurus was only 2.59 times larger in volume.[67]
Pathologies and health
In 2003, evidence of tumors, including hemangiomas, desmoplastic fibroma, metastatic cancer, and osteoblastoma, was described in Edmontosaurus bones. Rothschild et al. tested dinosaur vertebrae for tumors using computerized tomography and fluoroscope screening. Several other hadrosaurids, including Brachylophosaurus, Gilmoreosaurus, and Bactrosaurus, also tested positive. Although more than 10,000 fossils were examined in this manner, the tumors were limited to Edmontosaurus and closely related genera. The tumors may have been caused by environmental factors or genetic propensity.[69]
Osteochondrosis, or surficial pits in bone at places where bones articulate, is also known in Edmontosaurus. This condition, resulting from cartilage failing to be replaced by bone during growth, was found to be present in 2.2% of 224 edmontosaur toe bones. The underlying cause of the condition is unknown. Genetic predisposition, trauma, feeding intensity, alterations in blood supply, excess thyroid hormones, and deficiencies in various growth factors have been suggested. Among dinosaurs, osteochondrosis (like tumors) is most commonly found in hadrosaurids.[70]
Locomotion

Like other hadrosaurids, Edmontosaurus is thought to have been a facultative biped, meaning that it mostly moved on four legs, but could adopt a bipedal stance when needed. It probably went on all fours when standing still or moving slowly, and switched to using the hind legs alone when moving more rapidly.[4] Research conducted by computer modeling in 2007 suggests that Edmontosaurus could run at high speeds, perhaps up to 45 kilometres per hour (28 mph).[33] Further simulations using a subadult specimen estimated as weighing 715 kilograms (1,576 lb) when alive produced a model that could run or hop bipedally, use a trot, pace, or single foot symmetric quadrupedal gait, or move at a gallop. The researchers found to their surprise that the fastest gait was kangaroo-like hopping (maximum simulated speed of 17.3 metres per second (62 km/h; 39 mph)), which they regarded as unlikely based on the size of the animal and lack of hopping footprints in the fossil record, and instead interpreted the result as indicative of an inaccuracy in their simulation. The fastest non-hopping gaits were galloping (maximum simulated speed of 15.7 metres per second (57 km/h; 35 mph)) and running bipedally (maximum simulated speed of 14.0 metres per second (50 km/h; 31 mph)). They found weak support for bipedal running as the most likely option for high-speed movement, but did not rule out high-speed quadrupedal movement.[71]
While long thought to have been aquatic or semiaquatic, hadrosaurids were not as well-suited for swimming as other dinosaurs (particularly theropods, who were once thought to have been unable to pursue hadrosaurids into water). Hadrosaurids had slim hands with short fingers, making their forelimbs ineffective for propulsion, and the tail was also not useful for propulsion because of the ossified tendons that increased its rigidity, and the poorly developed attachment points for muscles that would have moved the tail from side to side.[72][73]
Social behavior
Extensive bone beds are known for Edmontosaurus, and such groupings of hadrosaurids are used to suggest that they were gregarious, living in groups.[4] Three quarries containing Edmontosaurus remains are identified in a 2007 database of fossil bone beds, from Alberta (Horseshoe Canyon Formation), South Dakota (Hell Creek Formation), and Wyoming (Lance Formation). One edmontosaur bone bed, from claystone and mudstone of the Lance Formation in eastern Wyoming, covers more than a square kilometre, although Edmontosaurus bones are most concentrated in a 40 hectares (0.15 sq mi) subsection of this site. It is estimated that disassociated remains pertaining to 10,000 to 25,000 edmontosaurs are present here. [74]
Unlike many other hadrosaurids, Edmontosaurus lacked a bony crest. It may have had soft-tissue display structures in the skull, though: the bones around the nasal openings had deep indentations surrounding the openings, and this pair of recesses are postulated to have held inflatable air sacs, perhaps allowing for both visual and auditory signaling.[35] Edmontosaurus may have been dimorphic, with more robust and more lightly built forms, but it has not been established if this is related to sexual dimorphism.[75]
Edmontosaurus has been considered a possibly migratory hadrosaurid by some authors. A 2008 review of dinosaur migration studies by Phil R. Bell and Eric Snively proposed that E. regalis was capable of an annual 2,600 kilometres (1,600 mi) round-trip journey, provided it had the requisite metabolism and fat deposition rates. Such a trip would have required speeds of about 2 to 10 kilometres per hour (1 to 6 mph), and could have brought it from Alaska to Alberta.[76][77] In contrast to Bell and Snively, Anusuya Chinsamy and colleagues concluded from a study of bone microstructure that polar Edmontosaurus overwintered.[78]
Chewing

Between the mid-1980s and the first decade of the 2000s, the prevailing interpretation of how hadrosaurids processed their food followed the model put forward in 1984 by David B. Weishampel. He proposed that the structure of the skull permitted motion between bones that resulted in backward and forward motion of the lower jaw, and outward bowing of the tooth-bearing bones of the upper jaw when the mouth was closed. The teeth of the upper jaw would grind against the teeth of the lower jaw like rasps, processing plant material trapped between them.[4][79] Such a motion would parallel the effects of mastication in mammals, although accomplishing the effects in a completely different way.[80] Work in the early 2000s has challenged the Weishampel model. A study published in 2008 by Casey Holliday and Lawrence Witmer found that ornithopods like Edmontosaurus lacked the types of skull joints seen in those modern animals that are known to have kinetic skulls (skulls that permit motion between their constituent bones), such as squamates and birds. They proposed that joints that had been interpreted as permitting movement in dinosaur skulls were actually cartilaginous growth zones.[81] An important piece of evidence for Weishampel's model is the orientation of scratches on the teeth, showing the direction of jaw action. Other movements could produce similar scratches though, such as movement of the bones of the two halves of the lower jaw. Not all models have been scrutinized under present techniques.[81] Vincent Williams and colleagues (2009) published additional work on hadrosaurid tooth microwear. They found four classes of scratches on Edmontosaurus teeth. The most common class was interpreted as resulting from an oblique motion, not a simple up-down or front-back motion, which is consistent with the Weishampel model. This motion is thought to have been the primary motion for grinding food. Two scratch classes were interpreted as resulting from forward or backward movement of the jaws. The other class was variable and probably resulted from opening the jaws. The combination of movements is more complex than had been previously predicted.[82]

Weishampel developed his model with the aid of a computer simulation. Natalia Rybczynski and colleagues have updated this work with a much more sophisticated three-dimensional animation model, scanning a skull of E. regalis with lasers. They were able to replicate the proposed motion with their model, although they found that additional secondary movements between other bones were required, with maximum separations of 1.3 to 1.4 centimetres (0.51 to 0.55 in) between some bones during the chewing cycle. Rybczynski and colleagues were not convinced that the Weishampel model is viable, but noted that they have several improvements to implement to their animation. Planned improvements include incorporating soft tissue and tooth wear marks and scratches, which should better constrain movements. They note that there are several other hypotheses to test as well.[80] Further research published in 2012 by Robin Cuthbertson and colleagues found the motions required for Weishampel's model to be unlikely, and favored a model in which movements of the lower jaw produced grinding action. The lower jaw's joint with the upper jaw would permit anterior–posterior motion along with the usual rotation, and the anterior joint of the two halves of the lower jaw would also permit motion; in combination, the two halves of the lower jaw could move slightly back and forth as well as rotating slightly along their long axes. These motions would account for the observed tooth wear and a more solidly constructed skull than modeled by Weishampel.[83]
Paleoecology
Distribution
Edmontosaurus was a wide-ranging genus in both time and space. The rock units from which it is known can be divided into two groups by age: the older Horseshoe Canyon and St. Mary River formations, and the younger Frenchman, Hell Creek, and Lance formations. The time span covered by the Horseshoe Canyon Formation and equivalents is also known as Edmontonian, and the time span covered by the younger units is also known as Lancian. The Edmontonian and Lancian time intervals had distinct dinosaur faunas.[84]

The Edmontonian land vertebrate age is defined by the first appearance of Edmontosaurus regalis in the fossil record.[85] Although sometimes reported as of exclusively early Maastrichtian age,[41] the Horseshoe Canyon Formation was of somewhat longer duration. Deposition began approximately 73 million years ago, in the late Campanian, and ended between 68.0 and 67.6 million years ago.[86] Edmontosaurus regalis is known from the lowest of five units within the Horseshoe Canyon Formation, but is absent from at least the second to the top.[87] As many as three quarters of the dinosaur specimens from badlands near Drumheller, Alberta may pertain to Edmontosaurus.[88]
Ecosystem
The Lancian time interval was the last interval before the Cretaceous–Paleogene extinction event that eliminated non-avian dinosaurs. Edmontosaurus was one of the more common dinosaurs of the interval. Robert Bakker reports that it made up one-seventh of the large dinosaur sample, with most of the rest (five-sixths) made up of the horned dinosaur Triceratops.[89] The coastal plain Triceratops–Edmontosaurus association, dominated by Triceratops, extended from Colorado to Saskatchewan.[90]

The Lance Formation, as typified by exposures approximately 100 kilometres (62 mi) north of Fort Laramie in eastern Wyoming, has been interpreted as a bayou setting similar to the Louisiana coastal plain. It was closer to a large delta than the Hell Creek Formation depositional setting to the north and received much more sediment. Tropical araucarian conifers and palm trees dotted the hardwood forests, differentiating the flora from the northern coastal plain.[91] The climate was humid and subtropical, with conifers, palmettos, and ferns in the swamps, and conifers, ash, live oak, and shrubs in the forests.[37] Freshwater fish, salamanders, turtles, diverse lizards, snakes, shorebirds, and small mammals lived alongside the dinosaurs. Small dinosaurs are not known in as great of abundance here as in the Hell Creek rocks, but Thescelosaurus once again seems to have been relatively common. Triceratops is known from many skulls, which tend to be somewhat smaller than those of more northern individuals. The Lance Formation is the setting of two edmontosaur "mummies".[91]
Diet

As a hadrosaurid, Edmontosaurus was a large terrestrial herbivore. Its teeth were continually replaced and packed into dental batteries that contained hundreds of teeth, only a relative handful of which were in use at any time.[4] It used its broad beak to cut loose food, perhaps by cropping,[4] or by closing the jaws in a clamshell-like manner over twigs and branches and then stripping off the more nutritious leaves and shoots.[53] Because the tooth rows are deeply indented from the outside of the jaws, and because of other anatomical details, it is inferred that Edmontosaurus and most other ornithischians had cheek-like structures, muscular or non-muscular. The function of the cheeks was to retain food in the mouth.[92][93] The animal's feeding range would have been from ground level to around 4 metres (13 ft) above.[4]
Before the 1960s and 1970s, the prevailing interpretation of hadrosaurids like Edmontosaurus was that they were aquatic and fed on aquatic plants.[94] An example of this is William Morris's 1970 interpretation of an edmontosaur skull with nonbony beak remnants. He proposed that the animal had a diet much like that of some modern ducks, filtering plants and aquatic invertebrates like mollusks and crustaceans from the water and discharging water via V-shaped furrows along the inner face of the upper beak.[47] This interpretation of the beak has been rejected, as the furrows and ridges are more like those of herbivorous turtle beaks than the flexible structures seen in filter-feeding birds.[94]
Because scratches dominate the microwear texture of the teeth, Williams et al. suggested Edmontosaurus was a grazer instead of a browser, which would be predicted to have fewer scratches due to eating less abrasive materials. Candidates for ingested abrasives include silica-rich plants like horsetails and soil that was accidentally ingested due to feeding at ground level.[82] The tooth structure indicates combined slicing and grinding capabilities.[52]
Reports of gastroliths, or stomach stones, in the hadrosaurid Claosaurus are actually based on a probable double misidentification. First, the specimen is actually of Edmontosaurus annectens. Barnum Brown, who discovered the specimen in 1900, referred to it as Claosaurus because E. annectens was thought to be a species of Claosaurus at the time. Additionally, it is more likely that the supposed gastroliths represent gravel washed in during burial.[5]
Gut contents
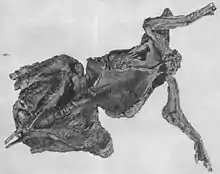
Both of the "mummy" specimens collected by the Sternbergs were reported to have had possible gut contents. Charles H. Sternberg reported the presence of carbonized gut contents in the American Museum of Natural History specimen,[95] but this material has not been described.[96] The plant remains in the Senckenberg Museum specimen have been described, but have proven difficult to interpret. The plants found in the carcass included needles of the conifer Cunninghamites elegans, twigs from conifer and broadleaf trees, and numerous small seeds or fruits.[97] Upon their description in 1922, they were the subject of a debate in the German-language journal Paläontologische Zeitschrift. Kräusel, who described the material, interpreted it as the gut contents of the animal,[97] while Abel could not rule out that the plants had been washed into the carcass after death.[98]
At the time, hadrosaurids were thought to have been aquatic animals, and Kräusel made a point of stating that the specimen did not rule out hadrosaurids eating water plants.[53][97] The discovery of possible gut contents made little impact in English-speaking circles, except for another brief mention of the aquatic-terrestrial dichotomy,[99] until it was brought up by John Ostrom in the course of an article reassessing the old interpretation of hadrosaurids as water-bound. Instead of trying to adapt the discovery to the aquatic model, he used it as a line of evidence that hadrosaurids were terrestrial herbivores.[53] While his interpretation of hadrosaurids as terrestrial animals has been generally accepted,[4] the Senckenberg plant fossils remain equivocal. Kenneth Carpenter has suggested that they may actually represent the gut contents of a starving animal, instead of a typical diet.[100][101] Other authors have noted that because the plant fossils were removed from their original context in the specimen and were heavily prepared, it is no longer possible to follow up on the original work, leaving open the possibility that the plants were washed-in debris.[96][102]
Isotopic studies

The diet and physiology of Edmontosaurus have been probed by using stable isotopes of carbon and oxygen as recorded in tooth enamel. When feeding, drinking, and breathing, animals take in carbon and oxygen, which become incorporated into bone. The isotopes of these two elements are determined by various internal and external factors, such as the type of plants being eaten, the physiology of the animal, salinity, and climate. If isotope ratios in fossils are not altered by fossilization and later changes, they can be studied for information about the original factors; warmblooded animals will have certain isotopic compositions compared to their surroundings, animals that eat certain types of plants or use certain digestive processes will have distinct isotopic compositions, and so on. Enamel is typically used because the structure of the mineral that forms enamel makes it the most resistant material to chemical change in the skeleton.[51]
A 2004 study by Kathryn Thomas and Sandra Carlson used teeth from the upper jaw of three individuals interpreted as a juvenile, a subadult, and an adult, recovered from a bone bed in the Hell Creek Formation of Corson County, South Dakota. In this study, successive teeth in columns in the edmontosaurs' dental batteries were sampled from multiple locations along each tooth using a microdrilling system. This sampling method takes advantage of the organization of hadrosaurid dental batteries to find variation in tooth isotopes over a period of time. From their work, it appears that edmontosaur teeth took less than about 0.65 years to form, slightly faster in younger edmontosaurs. The teeth of all three individuals appeared to show variation in oxygen isotope ratios that could correspond to warm/dry and cool/wet periods; Thomas and Carlson considered the possibility that the animals were migrating instead, but favored local seasonal variations because migration would have more likely led to ratio homogenization, as many animals migrate to stay within specific temperature ranges or near particular food sources.[51]
The edmontosaurs also showed enriched carbon isotope values, which for modern mammals would be interpreted as a mixed diet of C3 plants (most plants) and C4 plants (grasses); however, C4 plants were extremely rare in the Late Cretaceous if present at all. Thomas and Carlson put forward several factors that may have been operating, and found the most likely to include a diet heavy in gymnosperms, consuming salt-stressed plants from coastal areas adjacent to the Western Interior Seaway, and a physiological difference between dinosaurs and mammals that caused dinosaurs to form tissue with different carbon ratios than would be expected for mammals. A combination of factors is also possible.[51]
Predator-prey relationships

The time span and geographic range of Edmontosaurus overlapped with Tyrannosaurus, and an adult specimen of E. annectens on display in the Denver Museum of Nature and Science shows evidence of a theropod bite in the tail. Counting back from the hip, the thirteenth to seventeenth vertebrae have damaged spines consistent with an attack from the right rear of the animal. One spine has a portion sheared away, and the others are kinked; three have apparent tooth puncture marks. The top of the tail was at least 2.9 metres (9.5 ft) high, and the only theropod species known from the same rock formation that was tall enough to make such an attack is T. rex. The bones are partially healed, but the edmontosaur died before the traces of damage were completely obliterated. The damage also shows signs of bone infection. Kenneth Carpenter, who studied the specimen, noted that there also seems to be a healed fracture in the left hip which predated the attack because it was more fully healed. He suggested that the edmontosaur was a target because it may have been limping from this earlier injury. Because it survived the attack, Carpenter suggested that it may have outmaneuvered or outrun its attacker, or that the damage to its tail was incurred by the hadrosaurid using it as a weapon against the tyrannosaur.[103] However, more modern studies dispute the idea of an attack but rather other factors unrealated to an attack from a tyrannosaur.[104]
Another specimen of E. annectens, pertaining to a 7.6 metres (25 ft) long individual from South Dakota, shows evidence of tooth marks from small theropods on its lower jaws. Some of the marks are partially healed. Michael Triebold, informally reporting on the specimen, suggested a scenario where small theropods attacked the throat of the edmontosaur; the animal survived the initial attack but succumbed to its injuries shortly thereafter.[105] Some edmontosaur bone beds were sites of scavenging. Albertosaurus and Saurornitholestes tooth marks are common at one Alberta bone bed,[106] and Daspletosaurus fed on Edmontosaurus and fellow hadrosaurid Saurolophus at another Alberta site.[107]
See also
References
- Xing, H.; Mallon, J.C.; Currie, M.L. (2017). "Supplementary cranial description of the types of Edmontosaurus regalis (Ornithischia: Hadrosauridae), with comments on the phylogenetics and biogeography of Hadrosaurinae". PLOS ONE. 12 (4): e0175253. Bibcode:2017PLoSO..1275253X. doi:10.1371/journal.pone.0175253. PMC 5383305. PMID 28384240.
- Horner, John R.; Goodwin, Mark B.; Myhrvold, Nathan (2011). "Dinosaur Census Reveals Abundant Tyrannosaurus and Rare Ontogenetic Stages in the Upper Cretaceous Hell Creek Formation (Maastrichtian), Montana, USA". PLOS ONE. 6 (2): e16574. Bibcode:2011PLoSO...616574H. doi:10.1371/journal.pone.0016574. PMC 3036655. PMID 21347420.
- Bell, P. R.; Fanti, F.; Currie, P. J.; Arbour, V.M. (2013). "A Mummified Duck-Billed Dinosaur with a Soft-Tissue Cock's Comb". Current Biology. 24 (1): 70–75. doi:10.1016/j.cub.2013.11.008. PMID 24332547.
- Horner, John R.; Weishampel, David B.; Forster, Catherine A (2004). "Hadrosauridae". In Weishampel, David B.; Dodson, Peter; Osmólska Halszka (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 438–463. ISBN 978-0-520-24209-8.
- Creisler, Benjamin S. (2007). "Deciphering duckbills: a history in nomenclature". In Carpenter Kenneth (ed.). Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington and Indianapolis: Indiana University Press. pp. 185–210. ISBN 978-0-253-34817-3.
- Marsh, Othniel Charles (1892). "Notice of new reptiles from the Laramie Formation". American Journal of Science. 43 (257): 449–453. Bibcode:1892AmJS...43..449M. doi:10.2475/ajs.s3-43.257.449. S2CID 131291138.
- Marsh, Othniel Charles (1892). "Restorations of Claosaurus and Ceratosaurus". American Journal of Science. 44 (262): 343–349. Bibcode:1892AmJS...44..343M. doi:10.2475/ajs.s3-44.262.343. hdl:2027/hvd.32044107356040. S2CID 130216318. Archived from the original on 2018-07-22. Retrieved 2018-08-06.
- Lucas, Frederic A. (1904). "The dinosaur Trachodon annectens". Smithsonian Miscellaneous Collections. 45: 317–320.
- Hatcher, John B. (1902). "The genera and species of the Trachodontidae (Hadrosauridae, Claosauridae) Marsh". Annals of the Carnegie Museum. 1 (14): 377–386. Archived from the original on 2018-08-19. Retrieved 2014-04-08.
- Gilmore, Charles W. (1915). "On the genus Trachodon". Science. 41 (1061): 658–660. Bibcode:1915Sci....41..658G. doi:10.1126/science.41.1061.658. PMID 17747979.
- Norell, M. A.; Gaffney, E. S.; Dingus, L. (1995). Discovering Dinosaurs in the American Museum of Natural History. New York: Knopf. pp. 154–155. ISBN 978-0-679-43386-6.
- Dal Sasso, Cristiano; Brillante, Giuseppe (2004). Dinosaurs of Italy. Bloomington and Indianapolis: Indiana University Press. p. 112. ISBN 978-0-253-34514-1.
- Lambe, Lawrence M. (1902). "On Vertebrata of the mid-Cretaceous of the Northwest Territory. 2. New genera and species from the Belly River Series (mid-Cretaceous)". Contributions to Canadian Paleontology. 3: 25–81.
- Glut, Donald F. (1997). "Edmontosaurus". Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. pp. 389–396. ISBN 978-0-89950-917-4.
- Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. pp. 220–221.
- Weishampel, David B.; Horner, Jack R. (1990). "Hadrosauridae". In Weishampel, David B.; Dodson, Peter; Osmólska Halszka (eds.). The Dinosauria (1st ed.). Berkeley: University of California Press. pp. 534–561. ISBN 978-0-520-06727-1.
- Lambe, Lawrence M. (1917). "A new genus and species of crestless hadrosaur from the Edmonton Formation of Alberta" (pdf (entire volume, 18 mb)). The Ottawa Naturalist. 31 (7): 65–73. Archived from the original on 2018-08-19. Retrieved 2009-03-08.
- Campione, N.E.; Evans, D.C. (2011). "Cranial Growth and Variation in Edmontosaurs (Dinosauria: Hadrosauridae): Implications for Latest Cretaceous Megaherbivore Diversity in North America". PLOS ONE. 6 (9): e25186. Bibcode:2011PLoSO...625186C. doi:10.1371/journal.pone.0025186. PMC 3182183. PMID 21969872.
- Lambe, Lawrence M. (1920). The hadrosaur Edmontosaurus from the Upper Cretaceous of Alberta. Memoir. 120. Department of Mines, Geological Survey of Canada. pp. 1–79. ISBN 978-0-659-96553-0.
- Gilmore, Charles W. (1924). A new species of hadrosaurian dinosaur from the Edmonton Formation (Cretaceous) of Alberta. Bulletin. 38. Department of Mines, Geological Survey of Canada. pp. 13–26.
- Lambe, Lawrence M. (1913). "The manus in a specimen of Trachodon from the Edmonton Formation of Alberta". The Ottawa Naturalist. 27: 21–25.
- Lambe, Lawrence M. (1914). "On the fore-limb of a carnivorous dinosaur from the Belly River Formation of Alberta, and a new genus of Ceratopsia from the same horizon, with remarks on the integument of some Cretaceous herbivorous dinosaurs". The Ottawa Naturalist. 27: 129–135.
- Sternberg, Charles M. (1926). A new species of Thespesius from the Lance Formation of Saskatchewan. Bulletin. 44. Department of Mines, Geological Survey of Canada. pp. 77–84.
- Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. pp. 151–164.
- Glut, Donald F. (1982). The New Dinosaur Dictionary. Secaucus, NJ: Citadel Press. p. 57. ISBN 978-0-8065-0782-8.
- Brett-Surman, Michael K. (1989). A revision of the Hadrosauridae (Reptilia: Ornithischia) and their evolution during the Campanian and Maastrichtian. Ph.D. dissertation. Washington, D.C.: George Washington University.
- Brett-Surman, Michael K. (1975). The appendicular anatomy of hadrosaurian dinosaurs. M.A. thesis. Berkeley: University of California.
- Brett-Surman, Michael K. (1979). "Phylogeny and paleobiogeography of hadrosaurian dinosaurs" (PDF). Nature. 277 (5697): 560–562. Bibcode:1979Natur.277..560B. doi:10.1038/277560a0. S2CID 4332144. Archived (PDF) from the original on 2018-08-19. Retrieved 2018-08-06.
- Glut, Donald F. (1982). The New Dinosaur Dictionary. Secaucus, NJ: Citadel Press. pp. 49, 53. ISBN 978-0-8065-0782-8.
- Lambert, David; the Diagram Group (1983). A Field Guide to Dinosaurs. New York: Avon Books. pp. 156–161. ISBN 978-0-380-83519-5.
- Chapman, Ralph E.; Brett-Surman, Michael K. (1990). "Morphometric observations on hadrosaurid ornithopods". In Carpenter, Kenneth; Currie Philip J. (eds.). Dinosaur Systematics: Perspectives and Approaches. Cambridge: Cambridge University Press. pp. 163–177. ISBN 978-0-521-43810-0.
- Olshevsky, George. (1991). A Revision of the Parainfraclass Archosauria Cope, 1869, Excluding the Advanced Crocodylia. Mesozoic Meanderings No. 2. San Diego: Publications Requiring Research.
- "Mummified Dinosaur Unveiled". National Geographic News. 2007-12-03. Archived from the original on 2012-10-31. Retrieved 2007-12-03.
- Lee, Christopher (2007-12-03). "Scientists Get Rare Look at Dinosaur Soft Tissue". Washington Post. Archived from the original on 2008-10-13. Retrieved 2007-12-03.
- Hopson, James A. (1975). "The evolution of cranial display structures in hadrosaurian dinosaurs". Paleobiology. 1 (1): 21–43. doi:10.1017/S0094837300002165.
- Campione, N.E. (2009). "Cranial variation in Edmontosaurus (Hadrosauridae) from the Late Cretaceous of North America." North American Paleontological Convention (NAPC 2009): Abstracts, p. 95a. PDF link Archived 2019-05-05 at the Wayback Machine
- Derstler, Kraig (1994). "Dinosaurs of the Lance Formation in eastern Wyoming". In Nelson Gerald E. (ed.). The Dinosaurs of Wyoming. Wyoming Geological Association Guidebook, 44th Annual Field Conference. Wyoming Geological Association. pp. 127–146.
- Mori, Hirotsugu; Druckenmiller, Patrick S.; Erickson, Gregory M. (2015). "A new Arctic hadrosaurid from the Prince Creek Formation (lower Maastrichtian) of northern Alaska". Acta Palaeontologica Polonica. 61. doi:10.4202/app.00152.2015.
- Takasaki, R.; Fiorillo, A. R.; Tykoski, R. S.; Kobayashi, Y. (2020). "Re-examination of the cranial osteology of the Arctic Alaskan hadrosaurine with implications for its taxonomic status". PLOS ONE. 15 (5): e0232410. Bibcode:2020PLoSO..1532410T. doi:10.1371/journal.pone.0232410. PMC 7202651. PMID 32374777.
- Davies, K.L., 1983, Hadrosaurian dinosaurs of Big Bend National Park [M.A. thesis]: Austin, University of Texas, 231 p.
- Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loueff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth M.P.; Noto, Christopher N. (2004). "Dinosaur distribution". In Weishampel, David B.; Dodson, Peter; Osmólska Halszka (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 517–606. ISBN 978-0-520-24209-8.
- Wagner, J.R. 2001. The hadrosaurian dinosaurs (Ornithischia: Hadrosauria) of Big Bend National Park, Brewster County, Texas, with implications for Late Cretaceous Paleozoogeography. M.Sc. thesis, Texas Tech University, Austin, Tex.
- Lehman, Thomas M.; Wick, Steven L.; Wagner, Jonathan R. (2016). "Hadrosaurian dinosaurs from the Maastrichtian Javelina Formation, Big Bend National Park, Texas". Journal of Paleontology. 90 (2): 1–24. doi:10.1017/jpa.2016.48. S2CID 133329640.
- Lull, Richard Swann; Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. Geological Society of America Special Paper 40. Geological Society of America. pp. 50–93.
- Lambert, David; the Diagram Group (1990). The Dinosaur Data Book. New York: Avon Books. p. 60. ISBN 978-0-380-75896-8.
- Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. p. 225.
- Morris, William J. (1970). "Hadrosaurian dinosaur bills — morphology and function". Contributions in Science (Los Angeles County Museum of Natural History). 193: 1–14.
- Cope, Edward D. (1883). "On the characters of the skull in the Hadrosauridae". Proceedings of the Philadelphia Academy of Natural Sciences. 35: 97–107.
- Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. pp. 110–117.
- Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. pp. 128–130.
- Stanton Thomas, Kathryn J.; Carlson, Sandra J. (2004). "Microscale δ18O and δ13C isotopic analysis of an ontogenetic series of the hadrosaurid dinosaur Edmontosaurus: implications for physiology and ecology". Palaeogeography, Palaeoclimatology, Palaeoecology. 206 (2004): 257–287. Bibcode:2004PPP...206..257S. doi:10.1016/j.palaeo.2004.01.007.
- Erickson, Gregory M.; Krick, Brandon A.; Hamilton, Matthew; Bourne, Gerald R.; Norell, Mark A.; Lilleodden, Erica; Sawyer, W. Gregory. (2012). "Complex dental structure and wear biomechanics in hadrosaurid dinosaurs". Science. 338 (6103): 98–101. Bibcode:2012Sci...338...98E. doi:10.1126/science.1224495. PMID 23042891. S2CID 35186603. Archived from the original on 2018-08-19. Retrieved 2018-08-06.
- Ostrom, John H. (1964). "A reconsideration of the paleoecology of the hadrosaurian dinosaurs". American Journal of Science. 262 (8): 975–997. Bibcode:1964AmJS..262..975O. doi:10.2475/ajs.262.8.975.
- Galton, Peter M. (1970). "The posture of hadrosaurian dinosaurs". Journal of Paleontology. 44 (3): 464–473.
- Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. pp. 98–110.
- Osborn, Henry Fairfield (1909). "The epidermis of an iguanodont dinosaur". Science. 29 (750): 793–795. Bibcode:1909Sci....29..793F. doi:10.1126/science.29.750.793. PMID 17787819.
- Osborn, Henry Fairfield (1912). "Integument of the iguanodont dinosaur Trachodon". Memoirs of the American Museum of Natural History. 1: 33–54. hdl:2246/49.
- Manning, Phillip L.; Morris, Peter M.; McMahon, Adam; Jones, Emrys; Gize, Andy; Macquaker, Joe H. S.; Wolff, G.; Thompson, Anu; Marshall, Jim; Taylor, Kevin G.; Lyson, Tyler; Gaskell, Simon; Reamtong, Onrapak; Sellers, William I.; van Dongen, Bart E.; Buckley, Mike; and Wogelius, Roy A. (2009). "Mineralized soft-tissue structure and chemistry in a mummified hadrosaur from the Hell Creek Formation, North Dakota (USA)". Proceedings of the Royal Society B. 276 (1672): 3429–3437. doi:10.1098/rspb.2009.0812. PMC 2817188. PMID 19570788.CS1 maint: uses authors parameter (link)
- "Enough with the "Duck-Billed Dinosaurs"". Archived from the original on 2018-04-18. Retrieved 2018-04-18.
- "Shovel-Beaked, Not Duck-Billed". Archived from the original on 2018-04-18. Retrieved 2018-04-18.
- J, Morris, William (1970). "Hadrosaurian dinosaur bills — morphology and function". Contributions in Science. 193. ISSN 0459-8113. Archived from the original on 2018-08-19. Retrieved 2018-04-19.
- Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. p. 48.
- Gates, Terry A.; Sampson, Scott D. (2007). "A new species of Gryposaurus (Dinosauria: Hadrosauridae) from the late Campanian Kaiparowits Formation, southern Utah, USA". Zoological Journal of the Linnean Society. 151 (2): 351–376. doi:10.1111/j.1096-3642.2007.00349.x.
- Marsh, Othniel Charles (1893). "The skull and brain of Claosaurus". American Journal of Science. 45 (265): 83–86. Bibcode:1893AmJS...45...83M. doi:10.2475/ajs.s3-45.265.83. S2CID 131740074.
- Marsh, Othniel Charles (1896). "The dinosaurs of North America". Sixteenth Annual report of the United States Geological Survey to the Secretary of the Interior, 1894–1895: Part 1. Washington, D.C.: U.S. Geological Survey. pp. 133–244. doi:10.3133/ar16_1. Retrieved 2009-03-08.
- Brown, Barnum (1914). "Anchiceratops, a new genus of horned dinosaurs from the Edmonton Cretaceous of Alberta, with discussion of the ceratopsian crest and the brain casts of Anchiceratops and Trachodon". Bulletin of the American Museum of Natural History. 33: 539–548.
- Lull, Richard Swann; and Wright, Nelda E. (1942). Hadrosaurian Dinosaurs of North America. pp. 122–128.
- Jerison, Harry J.; Horner, John R.; Horner, Celeste C (2001). "Dinosaur forebrains". Journal of Vertebrate Paleontology. 21 (3, Suppl): 64A. doi:10.1080/02724634.2001.10010852. S2CID 220414868.
- Rothschild, B.M.; Tanke, D. H.; Helbling II, M.; Martin, L.D. (2003). "Epidemiologic study of tumors in dinosaurs". Naturwissenschaften. 90 (11): 495–500. Bibcode:2003NW.....90..495R. doi:10.1007/s00114-003-0473-9. PMID 14610645. S2CID 13247222.
- Rothschild, Bruce; Tanke, Darren H. (2007). "Osteochondrosis is Late Cretaceous Hadrosauria". In Carpenter Kenneth (ed.). Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs. Bloomington and Indianapolis: Indiana University Press. pp. 171–183. ISBN 978-0-253-34817-3.
- Sellers, W. I.; Manning, P. L.; Lyson, T.; Stevens, K.; Margetts, L. (2009). "Virtual palaeontology: gait reconstruction of extinct vertebrates using high performance computing". Palaeontologia Electronica. 12 (3): unpaginated. Archived from the original on 2009-12-18. Retrieved 2009-12-13.
- Bakker, Robert T. (1986). "The case of the duckbill's hand". The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction. New York: William Morrow. pp. 146–159. ISBN 978-0-8217-2859-8.
- Brett-Surman, M. K. (1997). "Ornithopods". In James O. Farlow; M. K. Brett-Surman (eds.). The Complete Dinosaur. Bloomington: Indiana University Press. pp. 330–346. ISBN 978-0-253-33349-0.
- Chadwick, Arthur; Spencer, Lee; Turner, Larry (2006). "Preliminary depositional model for an Upper Cretaceous Edmontosaurus bonebed". Journal of Vertebrate Paleontology. 26 (3, suppl): 49A. doi:10.1080/02724634.2006.10010069. S2CID 220413406.
- Gould, Rebecca; Larson, Robb; Nellermoe, Ron (2003). "An allometric study comparing metatarsal IIs in Edmontosaurus from a low-diversity hadrosaur bone bed in Corson Co., SD". Journal of Vertebrate Paleontology. 23 (3, suppl): 56A–57A. doi:10.1080/02724634.2003.10010538. S2CID 220410105.
- Bell, Phil R.; Snively, E. (2008). "Polar dinosaurs on parade: a review of dinosaur migration". Alcheringa. 32 (3): 271–284. doi:10.1080/03115510802096101.
- Lloyd, Robin (2008-12-04). "Polar Dinosaurs Endured Cold Dark Winters". LiveScience.com. Imaginova. Retrieved 2008-12-11.
- Chinsamy, A.; Thomas, D. B.; Tumarkin-Deratzian, A. R.; Fiorillo, A. R. (2012). "Hadrosaurs were perennial polar residents". The Anatomical Record. online preprint (4): 610–614. doi:10.1002/ar.22428. PMID 22344791. S2CID 42235761.
- Weishampel, David B. (1984). Evolution in jaw mechanics in ornithopod dinosaurs. Advances in Anatomy, Embryology, and Cell Biology. Advances in Anatomy, Embryology, and Cell Biology 87. 87. Berlin; New York: Springer-Verlag. pp. 1–109. doi:10.1007/978-3-642-69533-9. ISBN 978-0-387-13114-6. ISSN 0301-5556. PMID 6464809. S2CID 12547312.
- Rybczynski, Natalia; Tirabasso, Alex; Bloskie, Paul; Cuthbertson, Robin; Holliday, Casey (2008). "A three-dimensional animation model of Edmontosaurus (Hadrosauridae) for testing chewing hypotheses". Palaeontologia Electronica. 11 (2): online publication. Archived from the original on 2008-10-05. Retrieved 2008-08-10.
- Holliday, Casey M.; Witmer, Lawrence M. (2008). "Cranial kinesis in dinosaurs: intracranial joints, protractor muscles, and their significance for cranial evolution and function in diapsids". Journal of Vertebrate Paleontology. 28 (4): 1073–1088. doi:10.1671/0272-4634-28.4.1073. S2CID 15142387.
- Williams, Vincent S.; Barrett, Paul M.; Purnell, Mark A. (2009). "Quantitative analysis of dental microwear in hadrosaurid dinosaurs, and the implications for hypotheses of jaw mechanics and feeding". Proceedings of the National Academy of Sciences. 106 (27): 11194–11199. Bibcode:2009PNAS..10611194W. doi:10.1073/pnas.0812631106. PMC 2708679. PMID 19564603.
- Cuthbertson, Robin S.; Tirabasso, Alex; Rybczynski, Natalia; Holmes, Robert B. (2012). "Kinetic limitations of intracranial joints in Brachylophosaurus canadensis and Edmontosaurus regalis (Dinosauria: Hadrosauridae), and their implications for the chewing mechanics of hadrosaurids". The Anatomical Record. 295 (6): 968–979. doi:10.1002/ar.22458. PMID 22488827. S2CID 33114056.
- Dodson, Peter (1996). The Horned Dinosaurs: A Natural History. Princeton: Princeton University Press. pp. 14–15. ISBN 978-0-691-05900-6.
- Sullivan, Robert M.; Lucas, Spencer G. (2006). "The Kirtlandian land-vertebrate "age" – faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of western North America" (pdf). In Lucas, Spencer G.; Sullivan, Robert M. (eds.). Late Cretaceous Vertebrates from the Western Interior. New Mexico Museum of Natural History and Science Bulletin. 35. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 7–29.
- Wu, X-C.; Brinkman, D.B.; Eberth, D.A.; Braman, D.R. (2007). "A new ceratopsid dinosaur (Ornithischia) from the uppermost Horseshoe Canyon Formation (upper Maastrichtian), Alberta, Canada". Canadian Journal of Earth Sciences. 44 (9): 1243–1265. Bibcode:2007CaJES..44.1243W. doi:10.1139/E07-011.
- Eberth, David A. (2002). "Review and comparison of Belly River Group and Edmonton Group stratigraphy and stratigraphic architecture in the southern Alberta Plains" (PDF). Canadian Society of Petroleum Geology Diamond Jubilee Convention, Programs and Abstracts. 117: (cd). Archived from the original (PDF) on 2007-11-13. Retrieved 2009-03-08.
- Russell, Dale A. (1989). An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press, Inc. pp. 170–171. ISBN 978-1-55971-038-1.
- Bakker, Robert T. (1986). The Dinosaur Heresies. p. 438.
- Lehman, Thomas M. (2001). "Late Cretaceous dinosaur provinciality". In Tanke, Darren; Carpenter, Kenneth (eds.). Mesozoic Vertebrate Life. Bloomington and Indianapolis: Indiana University Press. pp. 310–328. ISBN 978-0-253-33907-2.
- Russell, Dale A. (1989). An Odyssey in Time: Dinosaurs of North America. pp. 180–181.
- Galton, Peter M. (1973). "The cheeks of ornithischian dinosaurs". Lethaia. 6: 67–89. doi:10.1111/j.1502-3931.1973.tb00873.x.
- Fastovsky, D.E., and Smith, J.B. (2004). "Dinosaur paleoecology." The Dinosauria. pp. 614–626.
- Barrett, Paul M. (2005). "The diet of ostrich dinosaurs (Theropoda: Ornithomimosauria)". Palaeontology. 48 (2): 347–358. doi:10.1111/j.1475-4983.2005.00448.x.
- Sternberg, Charles H. (1909). "A new Trachodon from the Laramie Beds of Converse County, Wyoming". Science. 29 (749): 753–54. Bibcode:1909Sci....29..747G. doi:10.1126/science.29.749.747. PMID 17840008.
- Currie, Philip J.; Koppelhus, Eva B.; Muhammad, A. Fazal (1995). ""Stomach" contents of a hadrosaurid from the Dinosaur Park Formation (Campanian, Upper Cretaceous) of Alberta, Canada". In Sun Ailing; Wang Yuangqing (eds.). Sixth Symposium on Mesozoic Terrestrial Ecosystems and Biota, Short Papers. Beijing: China Ocean Press. pp. 111–114. ISBN 978-7-5027-3898-3.
- Kräusel, R. (1922). "Die Nahrung von Trachodon". Paläontologische Zeitschrift (in German). 4 (2–3): 80. doi:10.1007/BF03041547. S2CID 186239385.
- Abel, O. (1922). "Diskussion zu den Vorträgen R. Kräusel and F. Versluys". Paläontologische Zeitschrift (in German). 4 (2–3): 87. doi:10.1007/BF03041547. S2CID 186239385.
- Wieland, G. R. (1925). "Dinosaur feed". Science. 61 (1589): 601–603. Bibcode:1925Sci....61..601W. doi:10.1126/science.61.1589.601. PMID 17792714.
- Carpenter, Kenneth (1987). "Paleoecological significance of droughts during the Late Cretaceous of the Western Interior". In Currie, Philip J.; Koster, Emlyn H. (eds.). Fourth Symposium on Mesozoic Terrestrial Ecosystems, Drumheller, August 10–14, 1987. Occasional Paper of the Tyrrell Museum of Palaeontology. 3. Drumheller, Alberta: Royal Tyrrell Museum of Palaeontology. pp. 42–47. ISBN 978-0-7732-0047-0.
- Carpenter, Kenneth (2007). "How to make a fossil: part 2 – Dinosaur mummies and other soft tissue" (PDF). The Journal of Paleontological Sciences. online. Archived from the original (PDF) on 2009-08-23. Retrieved 2009-03-08.
- Tweet, Justin S.; Chin, Karen; Braman, Dennis R.; Murphy, Nate L. (2008). "Probable gut contents within a specimen of Brachylophosaurus canadensis (Dinosauria: Hadrosauridae) from the Upper Cretaceous Judith River Formation of Montana". PALAIOS. 23 (9): 624–635. Bibcode:2008Palai..23..624T. doi:10.2110/palo.2007.p07-044r. S2CID 131393649.
- Carpenter, Kenneth (1998). "Evidence of predatory behavior by theropod dinosaurs". Gaia. 15: 135–144. Archived from the original on 2007-11-17. Retrieved 2009-03-08. [not printed until 2000]
- Siviero, ART V.; Brand, Leonard R.; Cooper, Allen M.; Hayes, William K.; Rega, Elizabeth; Siviero, Bethania C.T. (2020). "Skeletal Trauma with Implications for Intratail Mobility in Edmontosaurus Annectens from a Monodominant Bonebed, Lance Formation (Maastrichtian), Wyoming USA". PALAIOS. 35 (4): 201–214. Bibcode:2020Palai..35..201S. doi:10.2110/palo.2019.079. S2CID 218503493.
- Campagna, Tony (2000). "The PT interview: Michael Triebold". Prehistoric Times. 40: 18–19.
- Jacobsen, Aase Roland; Ryan, Michael J. (2000). "Taphonomic aspects of theropod tooth-marked bones from an Edmontosaurus bone bed (Lower Maastrichtian), Alberta, Canada". Journal of Vertebrate Paleontology. 19 (3, suppl): 55A. doi:10.1080/02724634.1999.10011202.
- "City Site Was Dinosaur Dining Room". ScienceDaily. ScienceDaily. 2007-07-03. Archived from the original on 2009-01-02. Retrieved 2008-12-07.
External links
 Media related to Edmontosaurus at Wikimedia Commons
Media related to Edmontosaurus at Wikimedia Commons
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||



.jpg.webp)