Drzewica Formation
The Drzewica Formation (Also Called Drzewica Series/Seria Drzewicka or Serii Drzewiekiej and in older literature Brónow Series) is a geologic formation in Szydłowiec, Poland. It is Pliensbachian in age.[1][2][3] Vertebrate fossils have been uncovered from this formation, including dinosaur tracks.[1] The Drzewica Formation is part of the Depositional sequence IV-VII of the late lower Jurassic Polish Basin, with the IV showing the presence of local Alluvial deposits, with possible meandriform deposition origin, dominated in Jagodne and Szydłowiec, while delta system occurred through the zone of the modern Budki.[1] The sequence V shows a reduction of the erosion in the Zychorzyn borehole of the Drzewica Formation, showing changes on the extension of the marine facies, where upper deposits change from Alluvial to Deltaic-Seashore depositional settings.[1] VI-VII facies where recovered on the Brody-Lubienia borehole, with a lower part exposed on the village of Śmiłów that shows a small fall of the Sea level.[1] The stathigraphic setting of the dinosaur tracks reported from the formation suggest a Seashore or Deltaic barrier. Body fossils reported include bivalves, palynology, fossil trunks, roots. Trunks of coniferous wood, especially Cheirolepidiaceae and Araucariaceae trees show the occurrence of vast coniferous forests around the tracksite.[4][5] The association of forests and dinosaur megafauna on the Pliensbachian suggests also a colder and specially damp ecosystem. As many studies of the formation share, Drzewica shows in part to be a gigantic shore barrel, setting at the time where the Polish basin sea was at its lowest point.[1] Other related units are Fjerritslev or Gassum Formation (Danish Basin), lower Bagå Formation (Bornholm), upper Neringa Formation (Lithuania). Abandoned informal units in Poland: upper Sawêcin beds, Wieluñ series or Bronów series.[6]
| Drzewica Formation Stratigraphic range: Pliensbachian ~188–183 Ma [1] | |
|---|---|
 The Śmiłów Sandstone Quarry outcrop of the formation, the main and location of the major tracksite recovered on it | |
| Type | Geological formation |
| Unit of | Kamienna Group[1] |
| Underlies | |
| Overlies |
|
| Thickness | Up to ~100 m (330 ft)[1] |
| Lithology | |
| Primary | Siliciclastic sandstones |
| Other | Several types of heteroliths and mudstones |
| Location | |
| Region | Szydłowiec, Southern Poland |
| Country | |
| Extent | Polish Liassic Basin[1] |
| Type section | |
| Named for | Drzewica, Poland |
| Named by | Karaszewski (as an informal unit)[2] |
 Drzewica Formation (Poland) Major Outcrop Location | |
Description
The Drzewica Formation between Gowarczów and Rozwady, forms Sandstone complexes of +10 or so meters, usually uniformly formed, finely less often medium-grained, separated by Silt-sandstone blocks, sandstone-like places or Siltstones.[7] In these sediments there are worm tubules (Ichnofossils), Rhizoids and the remains of Charred plants, sometimes thin carbon inserts.[7] This section on the area of Gowarczów-Rozwady wasn't completely covered, but connects the appearance of the Drzewica Formation strata with several coeval Boreholes: Gowarczów (GW-1 and GW-2), Kuraszków (KP-1 and KP-4), Sielec (A2), Adamów (Al), Kraszków (MG-2), Gielniów (GG-1) and in a few exposures (quarries) from the vicinity of Kraszków and Gielniów.[7] On this region the most complete profile of this formation was obtained in the Sielec Borehole (A2), which was pierced by the geological structure of the outcrop zone, covering the middle and upper section of the Drzewica Formation with a thickness of 146.7 m. Locally, the thickness of this series is estimated at 175-180 m.[7]
Paleogeography
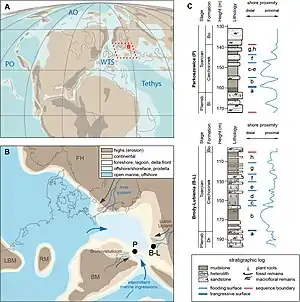
At the Late Pliensbachian (Margaritatus-Spinatum) several changes occur on the Polish Basin.[8] The last section, latest Raricostatum subzone, there were a series marine transgressive surfaces, linked with the called Pli1 transgressive phase, known thanks to molluscs and ooids, along with an increase of marine macrofauna and microfauna.[8] A marine Ingression filled the Polish basin, depositing marine dark Clay, Mudstone and lenticular Heteroliths, that are abundant on the lower-Middle Pliensbachian Łobez Formation and Poland, where dark-grey marine Claystones appear on the Kaszewy 1 borehole.[8] There was a visible Shoreline retreat that ends on the beginning of the Late Pliensbachian Margaritatus Chronozone.[8] After the Pli1, a series of local subordinate retrogradational stratal-pattern architectures indicate the transgressive phase of the sequence Pli2. Pli2 The transgressive phase led to a maximum flooding level at the middle Spinatum. That was known thanks to Ammonites found on an 85 m basin at NW Poland, on the Wolin IG-1 borehole.[9] This transgression also impacted deposition, with 265 m sequences of Mudstones and Sandstones in a syndepositional tectonic graben at the Wolin-Recław Zone.[10] After the maximun peak of the flooding basin it followed a regressive phase, characterized by progradational fluvio-deltaic Sandstones, like the series of Sandstones that recover the major tracksites of the Drzewica Formation, the Śmiłów Quarry Tracksite and the Wólka Karwicka Tracksite, both know from a regression nearshore/barrier lagoonar derived sandstone deposit.[10] The Coeval deposits of Usedom and East Prignitz on Germany contain numerous Glendonites (Calcite pseudomorphs after Ikaite), what lends support to the possibility of a Late Pliensbachian cooling event and polar Glaciation at that time.[11] Mostly of the authors, based on the current scientific quorum and the wide distribution of Glendonites in Upper Pliensbachian strata, tend to support a glacial-derived hypothesis.[12] The Spinatum regression collides with the worldwide Toarcian Oceanic Anoxic event. The oldest deposits of the sequence (Latest Pliensbachian-Lowermost Toarcian), partly of an age resting on a regional erosional surface, comprise alluvial and deltaic sediments assigned to a number of lithostratigraphic units of local significance.[8] The sequence recovers significant sea-level falls, that are related with the recurrent Late Pliensbachian glaciation, that would be followed at the start of the Toarcian with Global warming and a large scale trangression, recovered locally on the Ciechocinek Formation.[13]
Thanks to data from the Kaszewy 1 borehole levels of 1200 pCO2 were recovered, thanks to Plant stomatal index, being the highest amount recovered on the whole Lower Jurassic interval.[14] The studied strata also led to know a great variability in Total Organic Content, interpreted as result of a greater terrestrial biodegradation.[14] This led to knowing that the lower temperatures recovered from the Polish Basin in this interval occurred in the latest Rhaetian/earliest Hettangian, the late Hettangian, early Pliensbachian and especially in the Late Pliensbachian (Stokesi, Gibbosus, Spinatum), being this last the coolest period in the whole Early Jurassic.[14]
Stratygraphic profile
| Unit | Lithology | Thickness (metres) | Fossils |
|---|---|---|---|
|
Non Drzewica Formation (Ciechocinek Formation, Lower Toarcian)[15] |
Clay, Claystone |
2108,5–2111,0 m on the General Lithological Profile |
|
|
N1[15] |
Gray Sandstones |
2111.0–2115.0 m |
Non Reported |
|
N2[15] |
2115.0-2118.0 m |
Non Reported | |
|
N3[15] |
Sandstones, in the upper section salted, less clay downwards |
2118.0–2134.5 m |
Non Reported |
|
N4[15] |
Mudstone with sandstone inserts |
2134.5–2149.0 m |
Non Reported |
|
N5[15] |
Sandstone and infiltrated sandstone |
2149.0–2167.0 m |
Non Reported |
|
N6[15] |
Claystone |
2,167.0–2168.5 m |
Non Reported |
|
N7[15] |
Sandstones |
2168.5–2175.0 m |
Non Reported |
|
Core[15] |
|
2,175.0–2181.0+2.00 m |
|
|
SUB-Core (No Core)[15] |
Sandstones |
According to geophysical measurements up to a depth of 2178.0 m |
Non Reported |
Sedimentological evolution
The Formation strata starts on the Polish Jurassic Depositional sequence V, that besides Drzewica area and Budki–Szydłowiec–Jagodne area, is also known from the Brody-Lubienia borehole (with a massive exposure of the sequence V in an operating quarry near Bielowice, the major one along with Skrzynno near Przysucha), that is located on the eastern part of the Holy Cross Mts region, giving insight into development of Pliensbachian deposits in the marginal parts of this basin.[15] The initial part of this sequence is associated with a local fall of the sea level, where the lower IV level transition exposes with erosion, on locations such as the Zychorzyn borehole (at depth of 96.1 m) on Drzewica.[1] The latter sedimentation is marked with thin deposits from delta-distributary depositional subsystem, that become thicker around the Budki, where in the zone of the Brody-Lubienia borehole an alluvial-meandering channel deposition system developed.[1] After this initial regression there was a local transgression that led to abundance of local trace fossils and the formation of barrier-lagoonal deposits on Budki and in Brody-Lubienia, along with the increased presence of marine bivalves.[16][17] In the Brody-Lubienia borehole where developed in embayment-lagoon facies.[1] After the major marine transgression there was a visible change on Budki–Szydłowiec–Jagodne, shifting to fluvial discharge continuously moving along the Nowe Miasto–Iłża fault to the NE direction during the Pliensbachian times.[2] At this time on the Jagodne 1 borehole it was deposited the development of a barrier-lagoon linked with a delta.[2][18]
The Depositional sequence VI starts on the Jagodne 1 borehole with very large Mud clasts, pointing to intensive erosion of the underlying lagoonal sediments, while on the basin center, on the Zychorzyn borehole the erosion on lower sediments was lesser aggressive.[2] The sediments that where deposited at the start of this sequence are uniformly represented by coarse alluvial deposits, interpreted as meandering river with possible conspicuous share of bed load transport.[2] On the Budki 1 borehole series of coarse-grained alluvial package (especially large scale tabular cross-bedding sets dominate and quartz pebbles association), can represent low-sinuosity braided channels.[1] Another transgressive phase on this sequence led to th formation of foreshore/shoreface/barrier deposits, and a local event on the Polish Basin, where there was a rapid drowning of the palaeorelief with alluvial valleys and coastal plains, resulting in embayed coastline with detached beach/barrier ridges.[2] There was equilibrium with the sea level, with periodical little regressions and transgressions.[1][18] The maximun flooding range of this phase is represented by the Szydłowiec sandstones, developed on a regressive period where on Śmiłów was developed an eolian dune depositional subsystem linked with a barrier-lagoon, know due to the presence of barrier crest eolian deposits with plants buried. In this environment, the major Tracksites of the formation were developed.[1] Linked with the Śmiłów Quarry Sandstone appear deltaic deposits on the Brody-Lubienia borehole and on Zychorzyn.[1][18]
The Depositional sequence VII is the last Pliensbachian major sequence, know especially from the Brody-Lubienia borehole and the continuation of an alluvial portion exposed also at Śmiłów, where an exposure exhibits sequence VI/VII characters, showing that the sea level fall was not so significant, as it was in the base of the previous sequence.[1][18] This sequence starts with a rise of sea level and quick drowning of the whole area, destroying the previous delta system/marsh subsystem development measured at the Brody-Lubienia borehole, that shows that the amplitude of this transgression was quite high, as is represented by a thickness of 16 m.[1][18]
Śmiłów outcrop
There are several Types off Seashore Stratification seen on the Drzewica Formation. Starting with the post-Spinatum regression derived shore prograding cycle, where the nearshore deposits show detailed Hummocky cross-stratification, at the time are covered by thin beach-welded facies showing irregular “massive” bedding and numerous Plant roots penetrating down from the undulated bounding surface.[1] Superimposed are fine grained, very well sorted Sandstones with giant tabular Cross-bedding set interpreted as beach (Backshore depositional subsystem) Eolian Dune with complete Plant remains buried in the wind-transported Sand (Mostly, Herbaceous-Built Plants). The Direction of the Wind sedimentation is clearly visible on these deposits, where the stratification of the sand is well oriented. Those deposits are abundant on the Antecki quarry.[1] The most related to the shores, probably barrier-Lagoon and Delta depositional systems are clearly visible on the Smilów Quarry, where the exposure shows different lithofacies and strata, including fine-grained, white and light-grey Sandstones with Hummocky cross-stratification, with a series of tabular and trough Cross-bedding sets, probably related to nearmarine depositional subsystem.[1] A more fine-medium-grained, grey to brownish sandstones also with flat-tabular-horizontal bedding with abundant drifted Plant Fossils and plant remains in situ is considered to be part of a clear foreshoreb–arrier–Eolian depositional subsystem, probably related to a series of Dune fields, related to the Sea Barriers.[1] Kaolinized grey Mudstones with abundant Dispersed plant roots are related to more lagoonal depositional subsystems.[1] The slightly smaller and fine built, brownish and yellow Sandstones that show details Cross-bedding are related to a more alluvial deposition, probably from the influence of a local Channel.[1] On the Main Smilów Quarry the dominant shoreface depositional subsystem show detailed marks of clear Palaeocurrent directions, dispersed trough Cross-bedding inclinations that point to the West.[1] There have been measurements of the Wave-formed ripple that approximately reflects a shoreline orientation.[1] Thanks to the wave ripple crests orientation and the orientation of fossil tree logs it was indicated an inclination of the inclined tabular Foreset bed continuing across the whole outcrop.[1] These Foreset bed show a more pronounced inclination thanks to seaward-dipping Clinoforms, what reflects the original inclination of a barrier slope, with the sum of palaeocurrent pointing to south-west.[1]
CO2 Storage
Zaosie Anticline-Jeżów Anticline are two of the three structures selected as having the best conditions for geological CO2 storage in the Bełchatów region, with several wells drilled.[19] Both the structures are characterized by similar geological feature and petrophysical parameters of rocks, being potential reservoirs.[19] Significant thickness, good reservoir properties and proper depth give sufficient grounds to select the Lower Jurassic sandstone levels of the Drzewica Formation.[19]
Budziszewice-Zaosie structure is one of the potential CO2 storage sites for the biggest emitter of that gas in Europe, the Bełchatów Power Plant, with the major and most perspective reservoir located on the Drzewica Formation Strata locally.[20] Archival seismic sections subject to secondary interpretation made it possible for a more accurate mapping of the deep structure of the anticline and its definition as present state of recognition as tectonically continuous. The Major drilling hole was stablished on the top of the upper Pliensbachian, JPl3 (the top of the Drzewica formation of the first potential reservoir), with the (the top of the Ostrowiec formation as the second potential reservoir.[20] Parametric models of variability were made for the resulting geometrical matrix lithology, contamination, effective porosity and total permeability of the complexes.[20] Several works based on static 3D model recovered effective storage capacity c.a. 10 Mt (during 6 or 30 years of injection depending on hydrodynamic conditions).[20]
Fungi
Color key
|
Notes Uncertain or tentative taxa are in small text; |
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Saprophyte fungal Spores from Marine and Deltaic Settings associatted with wood and Litter.[21] The frequency of fungal spores on the Polish basin on the Pliensbachian-Toarcian transition is correlated with negative C isotope peaks and enhanced cuticular plant litter accumulation, pointing to climate-driven enhanced decomposition of wood and rapid destruction of terrestrial carbon, which may have played an important role in the aggravation of the Jurassic Greenhouse disaster.[21] Associated with a high rate of organic burial, the presence of Fungal Matter increased on the Uppermost layers of the Drzewica Formation, with a continued deposition between the T-OAE extinction, and several ups and downs on the Ciechocinek Formation, related with local climate and humidity changes. This is rather a reflection of the efficiency of terrestrial biodegradation.[21] Measured increasing of temperature favoured local fungal-mediated decomposition of plant litter, specifically of normally resistant wood.[21] Observed fungal spores represent various morphotypes and resemble superficially other palynogenic detritus, such as sphaerical Prasinophyceae.[21] In some cases spherical fungal spores show structures related to Hyphae attachments.[21] |
.jpg.webp) Spores found on the Polish are mostly from Saprophyte (wood decomposers) fungi, resembling the extant genera like Scutellinia, which are probably the main origin for the Fungal Spores found on the Ciechocinek Formation | |
|
Amerospores[21] |
|
|
Fungal Spores |
A Saprophyte fungus, member of the family Sordariomycetes inside Ascomycota.[21] Non-filamentous spores with no septations and with no projections longer than the spore body. Related with the extant genus Poronia. Fungal spore peaks linked to a relative and absolute loss of wood suggest a prominent role of fungal wood decomposers.[21] |
_Fr_587104.jpg.webp) Poronia punctata, an extant saprophyte fungus with Amerospores. Similar fungi probably liberated this spores on the Ciechocinek Formation |
|
Phragmospores[21] |
|
|
Fungal Spores |
A Saprophyte fungus, member of the family Dothideomycetes or Sordariomycetes inside Ascomycota.[21] Spores with two or more transverse septa. Related with the extant genus Acanthostigma, facultative pathogen, or beneficial partner of many plant species. Local humidity, though sufficiently high for fungal development throughout the whole T-OAE interval, is therefore thought to have been of a lesser significance for dynamics of fungal decomposition than elevated temperature.[21] |
 Leptosphaeria acuta, an extant wood fungus with phragmospores |
|
Dictyosporiaceae[21] |
|
|
Fungal Spores |
A freshwater anamorphic fungus, member of the family Pleosporales inside Pleosporomycetidae.[21] Multicellular spore with septations that intersect in more than one plane. Are characterized by being mostly aquatic lignicolous species with cheiroid, digitate, palmate and/or dictyosporous conidia. Related with the extant genus Dictyosporium, recorded worldwide from dead wood, decaying leaves, and palm material. |
 Lophiostoma compressum, an extant wood fungus with dictyospores |
Paleofauna

Along the Early Jurassic, there was a mostly marginal-marine (mainly Mesohaline), deltaic and continental sedimentation that prevailed in the area of Poland. A marine basin of Polyhaline character conditions occurred mainly in the Pliensbachian, due to a flooding of the Polish Basin.[22] Due to that, Pliensbachian sedimentation in Pomerania developed in a shelf basin that attained depths of about 100 m, below the storm wave base.[22] As the latest Pliensbachian began, the climate changes to a more cold environment, as prove the presence of ice sheets on northern latitude facies of the same age. That led to changes in salinity.[22] The Drzewica Formation was influenced by an ancient swampy-lagoonal sub-system, with Deltaic currents depositing fluvial sediments. The Main Boreholes show eustatic changes on the marine conditions, with a retreat of the sea level, leading to the appearance of vegetation and abundant pollen on the uppermost parts of the formation. The regressive phase of sea level is represented by the Komorowo Beds and Upper Slawecin Beds along with the own Drzewica Formation. As the level of the sea recovered on the Toarcian, most of the setting turn out to be a bathyal marine bottom.[23] The major outcrop of the formation, found on the Śmiłów Quarry represents a nearshore, deltaic, barrier/lagoonal ecosystem, where the biota left a high report of footprints (+200), with different sizes, that range from small to large animals of various groups. The Smilow Quarry Tracksite is the main outcrop and setting for the Footprints.[4] Along with the coeval (also belonging to the formation) Wólka Karwicka tracksite represent the only ones recovered from the Late Liassic of Europe.[4] The footprints are related to mostly lagoon-derived facies, unlike the vegetation, that appears on Deltaic facies. Brackish deposits are relates to Brachiopodans and other molluscs and Limuloid animals. The presence of Late Pliensbachian common ammonoids and several vertebrate clades connects the marine fauna with similar environments of the same age across Europe. The Drzewica Fauna is also, one of the few from the Pliensbachian-Toarcian Anoxic event recorded worldwide, specially referred to the vertebrate fauna reported.[24]
Ichnofossils
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Dwelling traces |
Marine Brackish or Freswather resting traces of Bivalves. The Lockeia traces trend to accumulate on Brackish waters on the case of the Drzewica Formation. They can occur in large aggregates, where occasionally traces present on agglomerations show a clear orientation, probably parallel to the palaeocurrent direction, which is associated with locomotion of the burrowing animal. |
 | |
|
|
Dwelling traces |
Marine Vertical funnel structure (simple or multiple) penetrated by a central straight or slightly curved (unbranched), plugged tube. Has been considered synonym of Skolithos. Fodichnnia related with marine worms (Arenicolidae) or worm-like organisms (Priapulida). |
 | |
|
|
Dwelling traces |
Marine, Brackish or Freshwater single, vertical, unbranched burrows, cylindrical or subcylindrical, lined or unlined, perfectly straight to curved, sometimes inclined to the vertical. Assigned to suspension-feeding worms (Arenicolidae, Spionida and Carpitellida) or Phoronidas. Alternatively, can be from Decapodans (Penaeidae and Anomura). |
 | |
|
|
Dwelling traces |
Marine, Brackish or Freshwater Unbranched U-shaped burrows having a subvertical orientation, with or without lining and passive fill. Are common on modern coastal environments, done by deposit-feeding polychaetes of the families Spionida and Carpitellida, as well as by suspension-feeding Amphipodan crustaceans and deposit-feeding Sipuncula. |
 | |
|
|
"U"-shaped burrows |
Marine-Mangroove Vertical, U-shaped, single-spreite Burrows; unidirectional or bidirectional spreite, generally continuous, rarely discontinuous.[26] Most Diplocraterion show only protrusive spreit, like the local ones, produced under predominantly erosive conditions where the organism was constantly burrowing deeper into the substrate as sediment was eroded from the top. "U"-shaped burrows, such as Diplocraterion, can be constructed by a wide variety of creatures: Polychaeta annelids (Axiothella, Abarenicola and Scolecolepis), Sipunculans (Sipunculus), Enteropneustans (Balanoglossus) and Echiurans (Urechis).[27] |
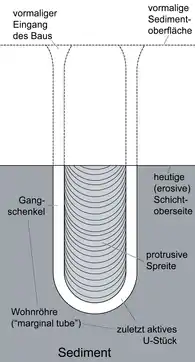 |
Bivalvia
Unidentified Blackish bivalves were reported from the formation on the first report of the Tracksites. Are related with shallow lagoonar facies.[4]
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Cunchs |
A Saltwater/Blackish Cockle, type member of the family Cardiniidae inside Carditida. On the Pliensbachian strata there is not a clear consensus if the Polish basin reached stenohaline conditions, but the presence of this genus on the Parkoszowice 58 BN borehole along with foraminiferans and marine phytoplankton tends to support more polyhaline-mesohaline conditions.[29] |
||
|
|
Cunchs |
A Saltwater/Blackish Clam, member of the family Arcticidae inside Cardiida. |
||
|
|
Cunchs |
A Saltwater/Blackish Clam, member of the family Tellinidae inside Cardiida. Thousands of individuals where found associated on a same level. |
.jpg.webp) Example of extant specimen of Macoma | |
|
|
Cunchs |
A Saltwater Clam, type member of the family Pleuromyidae inside Pholadida. A species are heretofore known solely from the Scania and Bornholm region, derived there from the Doshult and Katslosa series. |
||
|
|
Cunchs |
A Saltwater/Blackish venus Clam, member of the family Veneridae inside Cardiida. A lump was foun contained numerous fauna of clams assigned to the genus Cytherea. Is now believed that the liasic forms defined as Cytherea are of completely different Genera, probably Plesiocyprina. |
||
|
|
Cunchs |
A Saltwater/Blackish Clam, member of the family Corbiculidae inside Venerida. |
||
|
|
Cunchs |
A Saltwater/Blackish Pointed Nut Clam, type member of the family Nuculanidae inside Protobranchia. Originally described as Dacryomya zieteni. |
||
|
|
Cunchs |
A Freshwater Mussel, type member of the family Unionoida inside Palaeoheterodonta. The Only major Freshwater Bivalve recovered on the Formation. |
 Example of extant specimen of Unio | |
|
|
Cunchs |
A Freshwater Mussel, member of the family Unionidae inside Unionida. A great amount of shells were recovered along the deltaic facies of the Szydłowiec regression-derived sandstones, preserved mostly as imprints in the claystone. |
 Example of Unionidae Mussel |
Gastropoda
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Cunchs |
A Freshwater Snail, type member of the family Viviparidae inside Viviparoidea. In the area of the northern and north-western edge of the Swiętokrzyskie Mountains, the sea fauna has been found sporadically, but there are several finds of freshwater forms. Viviparus main identified snail on the Formation, and a typical form associated with Deltaic-Fluvial Deposits. |
 Example of extant specimen of Viviparus | |
|
|
Cunchs |
A Freshwater Snail, type member of the family Valvatidae inside Valvatoidea. Represented by small cunchs |
Ammonites
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Tragophylloceras loscombi |
|
MUZ.PGI.80.VI.149, single 1,7 cm Shell.[34] |
An Ammonite, member of the family Juraphyllitidae inside Ammonoidea. Latest Pliensbachian index fossil, Tragophylloceras is considered to be part of the Mediterranean-Submediterranean fossil region, but it has exceptions, with specimens coming from southern England and northwestern Germany.[34] In this case, the rocks that hosted the specimen also have poorly preserved dinoflagellate cysts, belonging to the genus Mendicodinium, that cannot expose its age beyond Late Pliensbachian.[34] This is supported by the presence of the fossil 20 m below the Lower Toarcian deposits. It looks also to be related to a large flooding surface, with the specimen being probably washed to nearshore or epicontinental waters.[34] |
 Example of
Tragophylloceras Fossils |
Crustacea
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Docens of Specimens |
A Brackish/Freshwater Clam shrimp (Phyllopodan) of the family Lioestheriidae. They fed on detritus, being very small slow moving, nektonic organisms that filter fed as they floated. The presence of Lioestheria marks the appearance of less saline conditions, as this is a mostly freshwater genus. The local Phyllopods are related with a great amount of freshwater debris (especially plants), and suggest seasonal changes in the rivers on the Pliensbachian-Toarcian Polish Basin. |
 Palaeestheria | |
|
Liasina[35] |
|
|
Valves |
A Brackish/Marine Ostracodan of the family Pontocyprididae. Small marine ostracods related with abundant Green Algae environments |
Limuloidea
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Limulitella cf. liasokeuperinus |
|
Head and partial upper body |
A marine/brackish Horseshoe crab, member of the family Xiphosura inside Xiphosurida (Protostomia). Only representative Limuloid from the Lower Jurassic, Limulitella cf. liasokeuperinus was found associated with brackish and deltatic facies, related to nearshore deposits.[37] Its affinities with the genus Limulitella has not been proved. It has been suggested as Limulitella incertade sedis, on the basis that there is not enough material to prove its affinities. The status of the species remain controversial, since the original fossil has been lost or in an unknown collection, more probably a private one.[39] |
||
|
Limulidae Indeterminate |
|
Unknown, only Cited |
A marine/brackish Horseshoe crab, member of the family Limulidae inside Xiphosurida. Body fossils reported from the Śmiłów Quarry tracksite include limuloid animals. Due to the environment representing a barrier/lagoonal setting, the specimens may have died during mating season. The specimens haven't been described. |
.jpg.webp) Limulus, example of Limuloid |
Chondrichthyes
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Szydłowiec sandstones |
Teeth |
A Freshwater/Marine Shark, member of the family Hybodontidae inside Hybodontiformes. Contemporaneous Hybodus fossils are known from Rumania. Due to the abundance of Hybodus on Lower Jurassic Strata this has been a temporal assignation. Beyond that, the specimen hasn't been compared. The genus resembles the freshwater sharks from coeval units of North America. |
 Restoration of Hybodus | |
|
Paleoxyris[40] |
|
Szydłowiec sandstones |
Egg Capsules |
A Freshwater/Brackish Shark Egg Capsule, probably from a member of the family of Lonchidiidae or Hybodontidae inside Hybodontiformes. With the sutdy of the contemporaneous flora from Chmielow (Gielniów Formation), Paleoxyris egg capsules where identified. On the same paper are claimed to be recovered also from the Szydłowiec sandstones.[40] This species is also known from the Rhaetian of Tübingen, Germany. The best candidate for this capsules can be the genus Lonchidion. |
Actinopteri
Unidentified fin spines are known from this formation.[4]
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Pholidophorus? angustus |
|
|
A marine Osteichthyes, type member of the family Pholidophoridae inside Teleostei. Possible Synonym of Furo angustus, was recovered from marine Ingression deposits, where thanks to marine pelecypods, was discovered that a brief marine ingression that took place on the area of the northern slopes of the Holy Cross Mountains in the final stage of the Lias (Late Pliensbachian-Toarcian). This genus lived on a basin with depths less than 50 m. |
 Pholidophorus | |
|
Semionotidae Indeterminate |
|
|
A Freshwater Osteichthyes, member of the family Semionotiformes inside Neopterygii. Fossils of Semionotus cf. bergeri are known from the Hettangian of Poland. The found teeth can be from a relative. Represents a genus similar to coeval fishes from inland deposits of North America. |
Sarcopterygii
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Cf. Coelacanthiformes sp. |
Szydłowiec sandstones |
|
A Freshwater Coelacanth, member of the family Coelacanthiformes inside Actinistia. The scales recovered are homologous to that found on the Triassic coelacanths Axelia & Wimania. Freshwater coelacanths are known from the Polish basin on the Triassic, especially from the Lipie Śląskie clay-pit at Lisowice. |
 Example of Freshwater Coelacanth (Chinlea) | |
|
Szydłowiec sandstones |
|
A Freshwater Lungfish, member of the Ptychoceratodontidae inside Ceratodontiformes. This Lugfish was originally assigned to the Triassic Polish genus Ceratodus silesiacus (Roemer, 1870), that has been known by several isolated tooth plates have been found in the lacustrine Triassic horizon of Krasiejów. But it resembles more the Lungfish fossils at the Triassic Lipie Śląskie clay-pit at Lisowice. Several Impressions of scales found in other Boreholes can belong also to Lugfish. It was found on Deltaic-Influenced deposits. |
 Example of Lugfish (Ceratodus) |
Rhynchocephalia
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Rhynchosauroides[43] |
|
|
Tracks |
Lacertoid tracks, member of the ichnofamily Rhynchosauroidae, incertade sedis inside Rhynchocephalia. Done by several Sphenodonts, such as Clevosaurus, Polysphenodon, and specially members of Gephyrosauridae, such as Gephyrosaurus and Deltadectes, also to Lepidosaurians, such as Squamatans (Relatives os the Jurassic basal squamatans, such as Scincomorpha members) . Lizard-built floor dwelling animals of different sizes, but rather small. |
 Rhynchosauroides Footprints were similar to the feet of the genus Gephyrosaurus |
Testudinata
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Ancyropus[4] |
|
|
Tracks |
Basal Turtle Ichnotaxon, member of the Testudines, incertade sedis inside Pantestudines. The relationships with Ancyropus are controversial, since the genus is known from the Early Liassic of North America. The tracks resemble Proganochelys, Palaeochersis, and probably other genera such as Kayentachelys or Australochelys. Small sized Turtles, with stimated less than 40 cm long, probably semiacuatic, related to the deltaic environments. Can be related to the Sinemurian and Toarcian Turtle fossils from Germany. |
Cynodontia
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Ameghinichnus[43] |
|
|
Tracks |
Cynodont tracks, type member of the ichnofamily Ameghinichnidae, incertade sedis inside Synapsida Represent tracks from Advanced Cynodonts, such as members of Tritylodontidae. The morphotypes resemble Oligokyphus, which is known from Pliensbachian Strata From England. Dinnebitodon, known from Pliensbachian strata of the Kayenta Formation, and larger ones to Bienotherium. That implicates the presence of various members of the late Cynodonts families, related to the modern mammals. Probably ground dwelling small animals that hunted insects or small vertebrates. |
 A genus similar to Oligokyphus is most probably the best candidate for the local Ameghinichnus footprints |
|
|
Tracks |
Represent tracks from Mammaliformes, member of the ichnofamily Chelichnopodidae, incertade sedis inside Synapsida. Resemble creatures such as Sinoconodon, Morganucodonta such as Erythrotherium, Megazostrodon or Dinnetherium; more advanced mammaliformes, such as members of Haramiyida, including arborear forms such as Shenshou. As show by Middle-Late Jurassic Finds, this group was ecologically diverse. |
 A genus similar to Morganucodon is most probably the best candidate for the local Brasilichnium footprints |
Crocodylomorpha
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Tracks |
|
_(20335774633).jpg.webp) Terrestrial crocodylomorphs such as Protosuchus, where most likely the Batrachopus trackmakers. |
Theropoda
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Footprints |
Theropod Tracks, member of the ichnofamily Anchisauripodidae, incertade sedis inside Theropoda. Considered a very Small Dinosaur, probably a juvenile. The Foot resembles those of young Coelophysis from Ghost Ranch Quarries.[45] Stenonyx footprints had been described from the Early Jurassic Soltykow (Hettangian) and Szydlowek, what matches with specimens found on the Hasle Formation, linking Southern Sweden, Bornholm and Poland, contiguous during the Early Jurassic and dinosaurs could thus freely roam this large area.[45] |
||
|
|
Footprints |
Theropod tracks, type member of the ichnofamily Anchisauripodidae, incertade sedis inside Neotheropoda. Assigned to Coelophysidae-alike dinosaurs. Small to medium slender primitive predatory dinosaurs, related with genera such as Liliensternus, Tachiraptor, Zupaysaurus, Procompsognathus or contemporaneous taxa such as the North American Segisaurus.[43][46] |
 Anchisauripus footprints belong to a genus similar to Procompsognathus | |
|
|
Footprints |
Theropod tracks, member of the ichnofamily Eubrontidae, incertade sedis inside Neotheropoda. The footprints from Smilow are small-sized,tridactyl and relatively narrow. forms Assigned to Coelophysidae-alike dinosaurs. Small to medium slender primitive predatory dinosaurs, related with neotheropods such as Camposaurus or Dracoraptor.[43][46] |
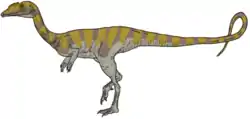 Grallator footprints belong to a genus similar to Camposaurus | |
|
|
Footprints |
Theropod tracks, member of the ichnofamily Eubrontidae, incertade sedis inside Neotheropoda. The tracks resemble Sinosaurus and are related with genera such as Notatesseraeraptor, Tachiraptor, Sarcosaurus and other basal Neotheropoda related to Tetanureae and Ceratosauriade. Small to medium-sized theropods. |
||
|
|
Footprints |
Theropod tracks, type member of the ichnofamily Eubrontidae, incertade sedis inside Theropoda. Eubrontes is related to the Genus Dilophosaurus, representing a basal Neotheropods. Include some large tracks, suggestings specimens up to 7 m, using Dilophosaurus as reference. Dilophosaurus is found or North America Pliensbachian strata, although the assignation of these tracks to the genus is controversial. Similar tracks have been described from Hettangian deposits from Poland and Sweden. There is a distinctive crouching theropod trace assigned to Eubrontes plexus, suggested a feather ingument on Dilophosauroids or relatives[25][48] |
 Model nicknamed "Dyzio", who represents a feathered reconstruction based on Drzewica and North American finds | |
|
Megalosauripus isp. |
|
Footprints |
Theropod tracks, member of the ichnofamily Eubrontidae, incertade sedis inside Theropoda. Largest Theropod Footprints from the Late Lower Jurassic. They have resemblance with the non-Tetanureae Sinosaurus, but match with Late Jurassic Orionides trackmakers. The trackmaker is expected to reach a length of 8.7-9.1 m long, with a weigth over 1.9-2.05 tonnes, making it among the largest Early Jurassic know theropod Trackmakers.[44] The footprints found on Drzewica seem even larger because of their large metatarsophalangeal area, which is often observed in the large theropod footprints from the post-Liassic strata.[44] |
 Megalosauripus footprints can belong to a large relative of Chuandongocoelurus |
Sauropodomorpha
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Footprints |
|
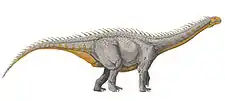 Local Parabrontopodus resemble the feet of the genus Barapasaurus |
Ornithischia
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Footprints |
Thyreophoran tracks, type member of the ichnofamily Moyenisauropodidae, incertade sedis inside Neornithischia. Is considered synonymous with the ichnogenus Anomoepus. The tracks adscribed resemble basal Thyreorphora foots. The tracks related to the genus are vinculated with genera such as Scelidosaurus, whose fossils have been found on Pliensbachian strata of England. In this case, implicates a ~4 m long animal. |
 Scelidosaurus matches with the bodyplan assigned to the Moyenisauropus trackmaker | |
|
|
Footprints |
Ornithischian tracks, member of the ichnofamily Moyenisauropodidae, incertade sedis inside Neornithischia. A foot similar to Lesothosaurus, implying 2.2 m long specimens. On the other casts there are tracks that resemble the feet of Stormbergia and various Genasauria of different sizes, between 90–130 cm. Fast-running small hervivorous ornithischians, probably gregarious. |
 Stormbergia matches with the bodyplan assigned to the Anomoepus trackmaker |
Plants
The sediments of this formation in Smiłów Quarry are mainly fine-grained Sandstones with small over-minerals. Diagonal layers and wave strata appear in them, where there is also traces of root plants with numerous plant remains. Juzuz Karaszewski observed in the Drzewica Sandstone vertically preserved plants, and on this basis he derived a possible dune-related origin of these sediments.[50] More recent studies suggest an environment of shallow seafront and beach, which of course does not exclude the presence of Aeolian zones there.[1] Lithofacies on the Smilów Quarry shows slightly inclined beach forests and abundant plant roots in a series of barrier/foreshore–ackshore–eolian dune facies, that include even plants buried in whole dunes, while drifted plant fossils and tree logs occur on the main depositional surface. This inclined surface exposes most likely a long period of non deposition and washed plants, recovering at the same time the original tilt of the local beach.[1]
Coals
The Kaszewy Coals found mostly on the Kaszewy-1 borehole and Niekłan PIG-1 borehole are the major coals of the Drzewica Formation and Ciechocinek Formation, representing the Pliensbachian-Toarcian boundary.[51] The Pliensbachian section of the Kaszewy-1 borehole comprises ~220 m of terrestrial and shallow marine siliciclastic sediments.[51] This Section was located on a nearshore-deltaic influenced setting, with an increased amount of both terrestrial and marine organic matter, reflecting increased weathering and increased transport of terrestrial organic matter, with ups and downs of the amount of both marine and terrestrial matter.[51] Here were collected fossil Charcoal in abundance and polycyclic aromatic hydrocarbon. Total fossil charcoal abundance is generally low in the Pliensbachian part of the study section, before peaking immediately above the Pl-To Boundary.[51] The abundance of coarse fossil charcoal particles (>125 μm) in the Pliensbachian-Toarcian sections of the Kaszewy-1 core is very low (0-15 particles/10 g sediment), while fine fossil charcoal abundance is generally low in the Pliensbachian level. There is also a greater abundance of non-charcoal particles.[51] At the start of the Toarcian locally, there is a measured increase on the amount of fine charcoal particles reflecting changes on the environment. Inside the Polycyclic Aromatic Hydrocarbons, the Pyrolytics (Benz-Anthracene, Benzo(k)Fluoranthene, Fluoranthene, Indeno[1,2,3-cd]Pyrene, Phenanthrene and Pyrene) where detected on a high variety of samples, with the Phenanthrene as the most abundant component, while Coronene is the lowest, suggesting an origin for this last one on the pyrolysis of organic matter.[51] Petrogenics are more abundant on the local coal samples than the Pyrolytic, what can suggest that these last ones may not reflect particularly local high levels of wildfire activity.[51] Apparently, the Kaszewy-1 borehole did not experience increased wildfire activity, but instead that the fine fossil charcoal abundance and Pyrolytics concentration records have showed a more clear regional wildfire signal.[51] The fossil charcoal abundance and geochemical data from Pliensbachian and Toarcian samples show evidence of at least four, and possible another two, levels of increased wildfire activity, and that these can be grouped into two larger periods of increased wildfire activity.[51] The increasing fires can be due to the recorded higher temperatures and decreased rainfall and humidity on the Late Pliensbachian, as result of the Global Cooling event.[52] This factors contribute to increasing the potential for wildfires to ignite and spread, and could explain the two levels of increased wildfire activity recorded in the Pliensbachian part of the study section.[51] The increased fire activity measured on the Pliensbachian level is related with peaks on the whole-plant C isotope discrimination local values, suggesting oxygen levels relatively high due to increased carbon burial.[53]
Palynology
The Drzewica Formation belongs to the Horstisporites planatus (Pl) Zone (Upper Sinemurian – Pliensbachian). This level starts on the older upper part of the Ostrowiec Formation, the Łobez Formation and Komorowo Formation of Pomerania and in other equivalent units from all Poland.[54] This Megaespore section of relatively poor compared with the lower and upper sections, with the lower boundary of the level is determined by the appearance of the indicator species Horisporites planatus (Selaginellaceae) and The upper limit of the level is marked by the appearance of continuous occurrences of Paxillitriletes phyllicus (Isöetaceae), indicating more humid conditions, and Erlansonisporites sparassis (Selaginellaceae).[54] This level has a series of characteristic megaespores, such as Minerisporites institus (Isöetaceae) and Hughesisporites pustulatus (Lycophyta) and on a minor ratio, rare occurrences of Hughesisporites planatus recorded even above the level range.[54] Is important to recover that the limit of the Pliensbachian-Toarcian in Poland was the first precisely defined on the basis of a detailed stratigraphic-sequential correlation, fully later confirmed by a high-resolution chemostratigraphic correlation, based on isotope analysis, with the ranges of individual megapores fitting well into this limit.[54] This level can be correlated with the non-megasporous level of Kuqaia quadrata, found in the sediments of the Sinemurian- Lower Pliensbachian Yangxia Formation in the Xinjiang Province, China.[54] In the Polish profiles, the Pl level correlates with the upper part of Rogalska´s 1st phase as well as the 2nd and 3rd microflora phase, where the following indicator myospores appear: Lycopodiumsporites gristhorpensis (Lycopodiaceae), Undulatisporites undulapolus (Filicopsida) and Applanopsipollenites segmentatus (Gymnospermopsida), and in central Poland represents the works of the Upper Sinemurian and the whole Pliensbachian, being probably correlated with the middle part of the myospore level Cerebropollenites macroverrucosus (Coniferophyta), distinguished in the Pliensbachian and Toarcian deposits in northern Europe.[54]
Megaflora
The Lublin Upland fluvial sandstones contain diverse types of fossil flora, associated genera and species only with Lower Jurassic sediments. Originally, while studying the Carboniferous flora from the boreholes in the area of the planned Bogdanka Coal Mine, appeared typical flora in similar to Jurassic formations.[55] The age of the plant material was not determined concretely until 2020, where was recovered as being Pliensbachian-Toarcian in age, with the flora representing an arid environment, more probably Pliensbachian, that is covered locally by the Drzewica Formation.[56] The Brody-Lubienia borehole is abundant on terrestrial Palynomorphs (know due to the presence of C29 Diasterenes, >70%), but also has high abundances of aquatic derived biomass.[56] Even with that, there is a clear consensus with the more terrestrial character of the sediments from Brody-Lubienia, expressed by the frequent occurrence of plant roots and paleosol horizons.[56] This also is recovered on therate of MTTCs (mono-, di- and tri-methyl-trimethyltridecyl-chromanes) where higher indices at Brody-Lubienia indicate lower salinities and a stronger influx of riverine freshwater.[56] The environment was probably Dry, developing flora on the near Freshwater influx settings. The Lublin Flora is linked with the flowing waters from the East.[56] Lublin lias is dominated by cycads and Bennetites Ginkgoales and pinnate, ferns occur sporadically, all on a conglomerate with numerous species occurs in the bottom of the Toarcian, where the deposits are filled with of coal, mudstone, sandstone and clay siderite (reworked from the Carboniferous), as well as pebbles from Devonian limestones.[57] Due to the fact that similar Boreholes and deposits nearby show similar characters it should be assumed that we have to dealing with the end of a river, that eroded the nearest Devonian-Carboniferous deposits (At the northeast), being brought by flowing waters and deposited in the aquatic environment-inland.[57] Vegetation mostly grew outside the sedimentation area, as well as on shores and shallows.[58]
| Genus | Species | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Cone fragment |
Affinities with Equisetidae inside Equisetopsida. Non-studied specimens of Horsetail Cones and where identified originally as Equisetum sp.. Are more probably related with the genus of equisetacean cones Equicalastrobus (specially Equicalastrobus chinleana), found on Eurasia during the late Triassic-Lower Jurassic Transition. It where described as resembling another fossil cone, Viridistachys. Cones from Herbaceous Equisetales, probably similar to the modern Equisetum.[59] |
||
|
|
Stems |
Affinities with Calamitaceae inside Equisetopsida. A common horsetail on the Liassic of Europe. Neocalamites lehmannianus was riginally described by Raciborski as Schizoneura hoerensis. |
 | |
|
|
Coiled Pinnae |
Incertae Ordinis inside Pteridophyta. Spiropteris is the name given to the fossil of a fern leaf before opening, when it is still coiled. |
||
|
|
Pinnae |
Affinities with the Peltaspermales inside Pteridospermatophyta. Bipinnate leaves, rachis longitudinally striated, with a long petiole and secondary rachises inserted oppositely to suboppositely on the upper side of the primary rachis. This Leaves belong to large Aboreal Ferns related with dry environments. |
||
|
|
Pinnae |
Affinities with the Umkomasiaceae inside Pteridospermatophyta. Is very similar to Rhaphidopteris, characterized by usually bi-tripinnate, coriaceous leaves with narrow segments. It belongs to Large (up to 25 m tall) arboreal Fern-like plants. |
||
|
|
Ovulate Structure |
Affinities with the Caytoniaceae inside Caytoniales. Represents the Ovulate organs or large Tree ferns, and is related with the middle Jurassic flora of United Kingdom. |
 | |
|
|
Pinnae |
Affinities with the Caytoniaceae inside Caytoniales. Sagenopteris is most likely the Pinnae of the Tree Fern that also contain Caytonia. |
 | |
|
|
Pinnae |
Affinities with the Bennettitales inside Bennettitopsida. Anomozamites is characterised by slender, (almost) completely and regularly segmented leaves whose leaflets are generally as long as broad or, at maximum, two times as long as broad. This genus is related with more arboreal Bennetitalean flora. Shows coriaceous leaves and is a genus linked more with dry climates. |
 | |
|
|
Pinnae |
Affinities with the Bennettitales inside Bennettitopsida. A more arbustive type of Bennetite, abundant on the Pliensbachian-Toarcian strata along Eurasia, related to rather dry climates. |
.jpg.webp) | |
|
|
Pinnae |
Affinities with the Bennettitales inside Bennettitopsida. This Bennetite has a leaf similar to Nilssonia, large, lanceolate in outline with coriaceous structure, like the modern angiosperm Philodendron. Is related with rather Dry-hot environments, with less Fern flora. |
||
|
|
Pinnae |
Affinities with the Cycadeoidaceae inside Bennettitales. The most abundant plant macrofossil locally, and the most diverse found on the Lublin coals. It represents a Cycad Like plant with Coriaceous leaves. Is related by some botanists with the Bennetite branch that led to Angiosperms. |
 | |
|
|
Bennetite "Flower" |
Affinities with the Cycadeoidaceae inside Bennettitales. The Bennetite flowers are the main organ that links this relatives with the Cycas with modern Angiosperms. Probably the Pollen assigned to Angiosperms come from plants with this "Flowers". |
||
|
|
Pinnae |
Affinities with the Williamsoniaceae inside Bennettitopsida. Leaves from Arboreal Bennetites, similar to the modern Cyca Encephalartos woodii, with robust Trunks, built for Dry and hot climates. |
 | |
|
|
|
Affinities with the Ginkgoaceae inside Ginkgoidae. Linked to the Hettangian-Sinemurian flora from Greenland and Skane, but also with coeval flora from the Sorthat Formation. Is the main Tree flora recovered locally. Is related with relatively warmer and humid conditions. It is the main recovered Floral cuticles on the Kasazewy 1 Borehole. |
 | |
|
|
Cuticles |
Affinities with the Ginkgoaceae inside Ginkgoidae. Is related with relatively warmer and humid conditions. |
 | |
|
|
Cuticles |
Affinities with the Ginkgoaceae inside Ginkgoidae. Is related with relatively warmer and humid conditions. |
||
|
Czekanowskia[14] |
|
|
Cuticles |
Affinities with the Czekanowskiales inside Ginkgoopsida. This Genus is related with relatively drier-cooler conditions. |
|
|
|
Cuticles |
Affinities with the Pseudotorelliaceae inside Ginkgoopsida. This Genus is related with relatively drier-cooler conditions. |
||
|
Swedenborgia?[64] |
|
|
MUZ PIG 76.III.1, Complete Plant |
Affinities with Krassiloviaceae inside Voltziales. A problematical plant that has been reported only on the Formation.[64] It is plant buried in whole in an eolian dune strata, composed mostly by Sandstone, that was deposited on a barrier-lagoon depositional system (possible backshore/backbarrier depositional subsystem).[64] With leaves of 5–25 cm long and about 2–5 mm wide, and an angle between them and the short main axis ranges within 20-30°, This taxon has a morphology that resembles Schizeacean ferns (Specially Schizaea pectinata), but also that of the Voltziales Aethophyllum stipulare from Anisian of France (A Herbaceous Conifer From the Triassic). It was proposed as an specimen of Swedenborgia on a revision of the specimens of the Hettangia from also Poland.[65] This suggests a herbaceous appearance of these plants, against traditional suppositions about their mainly tree-like character.[64] Finally, can be leaves similar to Podozamites (also Krassiloviaceae). If not herbaceous plants, it can be leaves from trees or shrubs, but all probably coming from an Aeolian-Dunar Environment.[64] |
|
|
|
Leaves |
Affinities with the Cheirolepidiaceae inside Pinales. The type Leave from the Cheirolepidaceae family (as Cheirolepis, the genus that give name to the family, is a junior synonym), appears to be linked with Hot climates, able to survive in dry, extreme conditions, and been fire tolerant. |
See also
References
- Pieñkowski, G. (2004). The epicontinental Lower Jurassic of Poland. Polish Geological Institute Special Papers, 12, 1-154.
- Karaszewski W., 1962 — The stratigraphy of the Lias in the Northern Mesozoic Zone surrounding the Święty Krzyż Mountains (Central Poland) [Eng. Sum.]. Pr. Inst. Geol., 30, cz. 3:333–416
- Złonkiewicz, Z., & Fijałkowska-Mader, A. (2018). Kamień w architekturze zespołu koœcielno-klasztornego na Karczówce w Kielcach (Góry Œwiętokrzyskie).
- Pienkowski, Grzegorz (2008). "Preliminary report about spectacular late Pliensbachian dinosaur tracksite from the Holy Cross Mountains, Poland. The 5th International Symposium of IGCP 506, Marine and non-marine Jurassic: global correlation and major geological events". 110111. Cite journal requires
|journal=(help) - Ziaja, J. (2006). Lower Jurassic spores and pollen grains from Odrowąż, Mesozoic margin of the Holy Cross Mountains, Poland. Acta Palaeobotanica, 46(1), 3-83.
- Gierliński, G. D., Ploch, I., Sabath, K., & Ziaja, J. (2006). Zagadkowa roślina z wczesnej jury Gór Świętokrzyskich. Przegląd Geologiczny, 54(2), 139-141.
- Cieśla, A. (1979). Budowa geologiczna strefy wychodni liasu między Gowarczowem a Rozwadami. Geological Quarterly, 23(2), 381-394.
- Barth, G., Pieńkowski, G., Zimmermann, J., Franz, M., & Kuhlmann, G. (2018). Palaeogeographical evolution of the Lower Jurassic: high-resolution biostratigraphy and sequence stratigraphy in the Central European Basin. Geological Society, London, Special Publications, 469(1), 341-369.
- Kopik, J. 1975. Fauna osadów domeru. In: Dembowska, J. (ed.) Wolin IG 1. Profile Głębokich Otworów Wiertniczych Instytutu Geologicznego, 22, 53.
- Dadlez, R. 1969. Stratigraphy of the Lias in Western Poland. Prace Instytutu Geologicznego, 57, 1–92 [in Polish with an English summary].
- Rogov, M.A. & Zakharov, V. 2010. Jurassic and Lower Cretaceous glendonite occurrences and their implication for Arctic paleoclimate reconstructions and stratigraphy. Earth Science Frontiers, 17, 345–346.
- Ruhl, M., Hesselbo, S.P. et al. 2017. Astronomical constraints on the duration of the Early Jurassic Pliensbachian Stage and global climatic fluctuations. Earth and Planetary Science Letters, 455, 149–165.
- Korte, C. & Hesselbo, S.P. 2011. Shallow marine carbon and oxygen isotope and elemental records indicate icehouse–greenhouse cycles during the Early Jurassic. Paleoceanography, 26, PA4219.
- Pieńkowski, G., Hesselbo, S. P., Barbacka, M., & Leng, M. J. (2020). Non-marine carbon-isotope stratigraphy of the Triassic-Jurassic transition in the Polish Basin and its relationships to organic carbon preservation, pCO2 and palaeotemperature. Earth-Science Reviews, 103383.
- Marek, S., KasińsKi, J., Krassowska, A., Leszczyński, K., Niemczycka, T., Feldman-Olszewska, A., ... & Gajewska, I. (2018). Szczegółowy profil litologiczno-stratygraficzny.
- Kopik J., 1962 — Faunistic criteria of stratigraphical subdivision of the Lias in North-Western and Central Poland [Eng. Sum.]. In: Ksiêga Pami¹tkowa ku czci prof. J. Samsonowicza (ed.E. Passendorfer): 271–312. Wyd. Geol. Warszawa
- Kopik J., 1964 — Stratigraphy of the Lower Jurassic based on the fauna of the Mechowo IG 1 borehole [Eng. Sum]. Biul. Inst. Geol., 189: 43–55.
- Pieńkowski, G. 1997. The Lower Jurassic: Sequence stratigraphy and sedimentology. In: S. Marek and M. Pajchlowa (Eds), The Epicontinental Permian and Mesozoic in Poland. Prace Państwowego Instytutu Geologicznego, 153, 217–235. [In Polish with English summary]
- Feldman-Olszewska, A., Adamczak, T., & Szewczyk, J. (2010). Charakterystyka poziomów zbiornikowych i uszczelniających w obrębie struktur Zaosia i Jeżowa pod kątem geologicznego składowania CO2 na podstawie danych z głębokich otworów wiertniczych. Biuletyn Państwowego Instytutu Geologicznego, (439 (1)), 17-28.
- Machowski W. , Papiernik B. (2010): Model dynamiczny zatłaczania CO2 do dolnojurajskich formacji solankowych struktury Budziszewice-Zaosie.Geopetrol 2010 : nowe metody i technologie zagospodarowania złóż i wydobycia węglowodorów w warunkach lądowych i morskich : międzynarodowa konferencja naukowo-techniczna : Zakopane, 20--23.09.2010
- Pieńkowski, G., Hodbod, M., & Ullmann, C. V. (2016). Fungal decomposition of terrestrial organic matter accelerated Early Jurassic climate warming. Scientific reports, 6, 31930.
- Pieñkowski, G., & Waksmundzka, M. (2009). Palynofacies in Lower Jurassic epicontinental deposits of Poland: tool to interpret sedimentary environments. Episodes, 32(1), 21-32.
- Feldman-Olszewska, A. (1997). Depositional systems and cyclicity in the intracratonic Early Jurassic basin in Poland. Geological Quarterly, 41(4), 475-490.
- BRAŃSKI, P. (2012). The mineralogical record of the Early Toarcian stepwise climate changes and other environmental variations (Ciechocinek Formation, Polish Basin). Volumina Jurassica, 10(10), 1-24.
- Gerard Dariusz Gierlinski, Martin G. Lockley, Grzegorz Niedźwiedzki. "A distinctive crouching theropod trace from the Lower Jurassic of Poland". Geological Quarterly. 53(4): 23 281.CS1 maint: uses authors parameter (link)
- Chakraborty, A., & Bhattacharya, H. N. (2013). Spreiten burrows: a model based study on Diplocraterion parallelum. Geospectrum, 296-299.
- Ekdale, A. A., & Lewis, D. W. (1991). Trace fossils and paleoenvironmental control of ichnofacies in a late Quaternary gravel and loess fan delta complex, New Zealand. Palaeogeography, Palaeoclimatology, Palaeoecology, 81(3-4), 253–279. doi:10.1016/0031-0182(91)90150-p
- Kopik, J. (1960). O kilku morskich małżach z serii gielniowskiej liasu Gór Świętokrzyskich [About several sea clams from the series of Gielniowska lias of the Świętokrzyskie Mountains]. Geological Quarterly , 4 (1), 95-104.
- Kilhams, B., Kukla, P. A., Mazur, S., McKie, T., Mijnlieff, H. F., van Ojik, K., & Rosendaal, E. (2018). Mesozoic resource potential in the Southern Permian Basin area: the geological key to exploiting remaining hydrocarbons whilst unlocking geothermal potential. Geological Society, London, Special Publications, 469(1), 1-18.
- Krajewski R. (1947) - Złoża żelaziaków ilastych we wschodniej części powiatu koneckiego. Biul. Państw. Inst. GeoL, 26. Warszawa.
- Ksiązkiewicz M., Samsonowicz J. (1952) - Zarys geolcigii Polski. PWN, p.90-130. Warszawa
- Samsonowicz J. (1929) - Cechsztyn, trias i lias na północnym obrżeżeniu Łysogór. Spraw. Państ. Inst. Geol, 5, nr 1-2, p. ;1-282. Warszawa.
- Samsonowicz J. (1934) - Objaśnienia arkusza Opatów. Ogólna mapa geologiczna Poląkiw skali 1: 100000. Państw. Inst. GeoL Warszawa
- Pienkowski, Grzegorz (2014): The first Early Jurassic ammonite find in central Poland. Volumina Jurassica, 2014, XII (1): 99–104:https://www.researchgate.net/publication/297380409_The_first_Early_Jurassic_ammonite_find_in_central_Poland
- Kopik, J. (1998). Lower and Middle Jurassic of the north-eastern margin of the Upper Silesian Coal Basin. Biuletyn Państwowego Instytutu Geologicznego, 378, 67-129.
- Kopik J. and Marcinkiewicz T. 1997. Jura dolna: Biostratygrafia. In: S. Marek and M. Pajchlowa (Eds), The epicontinental Permian and Mesozoic in Poland (in Polish with English summary). Prace Paƒstwowego Instytutu Geologicznego, 153: 196-205.
- Braun, C.F., 1860, Die Thiere in den Pflanzenschiefern der Gegend von Bayreuth: 1771, Zoologica Fundamenta
- Karaszewski, W. (1975). Obecność rzadkiej skamieniałości-limulusa w liasie świętokrzyskim. Przegląd Geologiczny, 23(10), 500.
- Błażejowski, B., Niedźwiedzki, G., Boukhalfa, K., & Soussi, M. (2017). Limulitella tejraensis, a new species of limulid (Chelicerata, Xiphosura) from the Middle Triassic of southern Tunisia (Saharan Platform). Journal of Paleontology, 91(5), 960-967.
- W. Karaszewski. 1965. O srodkowoliasowym wieku flory z Chmielowa pod Ostrowcem i jej znaczeniu dla stratygrafii kontynentalnej jury [On the Middle Liassic age of the flora from Chmielow, near Ostrowiec (central Poland) and its significance for stratigraphy of the continental Jurassic].
- Agassiz, L. (1844). Tableau général des poissons fossiles rangés par terrains. Imprimerie de Petitpierre.
- Kopik, J. (1960). Of the Lias in the Święty Krzyż Mountains. Kwartalnik geologiczny, 4(1), 103.
- Niedźwiedzki, G., Remin, Z., Roszkowska, J., & Meissner, U. (2008). New finds of dinosaur tracks in the Liassic strata of the Holy Cross Mountains, Poland. Przegląd Geologiczny, 57, 252-262.
- Grzegorz Niedźwiedzki, zbyszek Remink: Gigantic theropod dinosaur footprints from the Upper Pliensbachian of the Holy Cross Mountains, Poland [in Polish with English abstract].Article in Przeglad Geologiczny 56(9):823-825 · September 2008
- Jesper Milàn, Finn Surlyk. 2014: An enigmatic, diminutive theropod footprint in the shallow marine Pliensbachian Hasle Formation, Bornholm, Denmark
- Gierlinski, G. D. (2007). New dinosaur tracks in the Triassic, Jurassic and Cretaceous of Poland. Jornadas Internacionales sobre Paleontoloia de Dinosaurios y su Entrono, Salas de los Infantes, Burgos, Spain, 13-16.
- G. Gierlinski and G. Niedzwiedzki. 2005. New saurischian dinosaur footprints from the Lower Jurassic of Poland. Geological Quarterly 49(1):99-104
- Li, D. Q., Xing, L. D., Lockley, M. G., Romilio, A., Yang, J. T., & Li, L. F. (2019). The first theropod tracks from the Middle Jurassic of Gansu, Northwest China: new and rare evidence of quadrupedal progression in theropod dinosaurs. Journal of Palaeogeography, 8(1), 10.
- Gierliński, G. D. (2009). A preliminary report on new dinosaur tracks in the Triassic, Jurassic and Cretaceous of Poland. Actas de las IV Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno Salas de los In fan tes, Burgos, 75-90.
- Karaszewski W. & Kopik J. 1970 — Jura dolna. Inst. Geol. Pr., 56:65–98
- Pointer, R. (2019). Fire & Global Change During Key Intervals of the Late Triassic & Early Jurassic with a Focus on the Central Polish Basin.
- Bailey, T. R., Rosenthal, Y., McArthur, J. M., Van de Schootbrugge, B., & Thirlwall, M. F. (2003). Paleoceanographic changes of the Late Pliensbachian–Early Toarcian interval: a possible link to the genesis of an Oceanic Anoxic Event. Earth and Planetary Science Letters, 212(3-4), 307-320.
- Ikeda, M., Hori, R. S., Ikehara, M., Miyashita, R., Chino, M., & Yamada, K. (2018). Carbon cycle dynamics linked with Karoo-Ferrar volcanism and astronomical cycles during Pliensbachian-Toarcian (Early Jurassic). Global and planetary change, 170, 163-171.
- Marcinkiewicz, T., Fijałkowska-Mader, A., & Pieńkowski, G. (2014). Megaspore zones of the epicontinental Triassic and Jurassic deposits in Poland–overview. Biuletyn Państwowego. Instytutu Geologicznego, 457, 15-42.
- Migier T. 1978. Nowe stanowiska flory jurajskiej w Lubelskim Zagłębiu Węglowym. Materiały III Naukowej Konferencji Paleontologów poświęconej badaniom regionu górnośląskiego oraz karbonu LZW i GZW. Streszczenia komunikatów: 33–34.Uniwersytet Śląski, Katowice.
- Ruebsam, W., Pieńkowski, G., & Schwark, L. (2020). Toarcian climate and carbon cycle perturbations–its impact on sea-level changes, enhanced mobilization and oxidation of fossil organic matter. Earth and Planetary Science Letters, 546, 116417.
- Samsonowicz, J. (1929). Cechsztyn, trias i lias na północnem zboczu Łysogór.. PIG, Sprawozdania,t. V, z. 1-2
- Szydeł, Z., & Szydeł, R. (1981). Profil utworów liasu na obszarze Lubelskiego Zagłębia Węglowego. Przegląd Geologiczny, 29(11), 568-571.
- Raciborski, M. (1891). Flora retycka północnego stoku Gór Świętokrzyckich. Nakł. Akademii Umiejętności.
- RACIBORSKI, M. (1891). Ueber die rhätische Flora am Nordabhange des polnischen Mittelgebirges. Anz. Akad. Wissenschaften Krakau, 64-68.
- Pacyna, G. (2013). Critical review of research on the Lower Jurassic flora of Poland. Acta Palaeobotanica, 53(2), 141-163.
- MAKAREWICZÓWNA A. 1928. Flora dolno-liasowaokolic Ostrowca (summary: Étude sur la flore fossile du lias inférieur des environs d’Ostrowiec). Prace Towarzystwa Przyjaciół Nauk w Wilnie, Wydział Nauk Matematycznych i Przyrodniczych, Prace Zakładu Geologicznego Uniwersytetu St. Batorego w Wilnie (Travaux de la Société des Science et des Letters de Wilno, Travaux de L’Institut de Géologie de l’Université de Wilno), 4(3): 1–49.
- PUSCH G.G. 1833. Geologische Beschreibung von Polen so wie der übrigen Nordkarpathen-Länder. Erster Theil. J.G. Cotta’schen Buchhandlung, Stuttgart und Tübingen.
- Gierliñski, G.D., Ploch, I., Sabath, K., Ziaja, J., 2006. An Early Jurassic problematical plant from the Holy Cross Mountains, central Poland. Prz. Geol. 54, 139–141. https://www.researchgate.net/publication/289572652_An_Early_Jurassic_problematical_plant_from_the_Holy_Cross_Mountains_central_Poland
- Gierliński, G., Pieńkowski, G., & Niedźwiedzki, G. (2004). Tetrapod track assemblage in the Hettangian of Sołtyków, Poland, and its paleoenvironmental background. Ichnos, 11(3-4), 195-213.