Lobopodia
The lobopodians, members of the informal group Lobopodia[2] (from the Greek, meaning "blunt feet"), or the formally erected phylum Lobopoda Cavalier-Smith (1998),[3] are panarthropods with stubby legs called lobopods,[4] a term which may also be used as a common name of this group as well.[5][6] While the definition of lobopodians may differ between literatures, it usually refers to a group of soft-bodied, worm-like fossil panarthropods such as Aysheaia and Hallucigenia.[4]
| Lobopodia | |
|---|---|
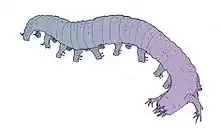 | |
| Reconstruction of the lobopodian Aysheaia pedunculata | |
| Scientific classification | |
| Kingdom: | Animalia |
| Subkingdom: | Eumetazoa |
| Clade: | ParaHoxozoa |
| Clade: | Bilateria |
| Clade: | Nephrozoa |
| (unranked): | Protostomia |
| Superphylum: | Ecdysozoa |
| (unranked): | Panarthropoda |
| Phylum: | †"Lobopodia" Snodgrass 1938 |
| Groups included | |
| |
| Cladistically included but traditionally excluded taxa | |
|
Crown-group Euarthropoda | |
| Synonyms | |
| |

The oldest near-complete fossil lobopodians date to the Lower Cambrian; some are also known from Ordovician, Silurian and Carboniferous Lagerstätten.[7][8][9] Some bear toughened claws, plates or spines, which are commonly preserved as carbonaceous or mineralized microfossils in Cambrian strata.[10][11]
Definition
The scope of the Lobopodian concept varies from author to author. Its most general as well as the most limited sense refers to a suite of mainly Cambrian worm-like panarthropod taxa with lobopods – for example Aysheaia, Hallucigenia and Xenusion, members which were traditionally united as "xenusians" or "xenusiids" (class Xenusia). The dinocaridid genera Pambdelurion and Kerygmachela may also be regarded as lobopodians,[12][13] eventually referred as "gilled lobopodians" or "gilled lobopod".[14] Under such definitions, "Lobopodia" compose of only extinct taxa, and widely accepted as an informal, paraphyletic grade in correspond to the crown-group of three extant panarthropod phyla: Onychophora (velvet worms), Tardigrada (waterbears) and Arthropoda (arthropods).[4][15][16][17][18][19]
An alternative, broader definition of lobopodians would also incorporate the extant phyla Onychophora and Tardigrada,[20][12][3] two groups of panarthropod which also bore lobopodous limbs as well.[4] "Lobopodia" may also refer to a possible clade sister to Arthropoda, and compose of only Tardigrada and Onychophora.[21] Lobopodia sometimes also included Pentastomida, a group of parasitic panarthropod which later revealed to be a highly specialized taxon of crustacean arthropods.[22][23] The broadest definition proposes the monophyletic superphylum Lobopodia is equivalent to Panarthropoda.[24][5]
Representative taxa


The better-known genera include, for example, Aysheaia, which was discovered in the Canadian Burgess Shale and Hallucigenia, known from both Chenjiang Maotianshan Shale and the Burgess Shale. Aysheaia pedunculata has morphology apparently basic for lobopodians[15] — for example significantly annulated cuticle, terminal mouth opening, specialized frontalmost appendages and stubby lobopods with terminal claws. Hallucigenia sparsa is famous by having a complex history of interpretation — it was originally reconstructed with long, stilt-like legs and mysterious fleshy dorsal protuberances, and was long considered a prime example of the way in which nature experimented with the most diverse and bizarre body designs during the Cambrian.[25] However, further discoveries showed that this reconstruction had placed the animal upside-down: interpreting the "stilts" as dorsal spines made it clear that the fleshy "dorsal" protuberances were actually elongated lobopods. More recent reconstruction even exchanged the front and rear ends of the animal: reveal the bulbous inprint previously thought to be a head was actually gut contents being expelled from its anus.[10][16]
Microdictyon is another charismatic as well as the speciose genus of lobopodians resembling Hallucigenia, but instead of spines, it bore pairs of net-like plates which often found disarticulated and known as an example of small shelly fossils (SSF). Xenusion has the oldest fossil record amongst described lobopodians which may trace back to Cambrian Stage 2.[20][13] Luolishania is an iconic example of lobopodians with multiple pairs of specialized appendages.[26] The gill lobopodians Kerygmachela and Pambdelurion shed light on the relationship between lobopodians and arthropods, as they have both lobopodian affinities and characters link to the arthropod stem-group.[14][24]
Morphology
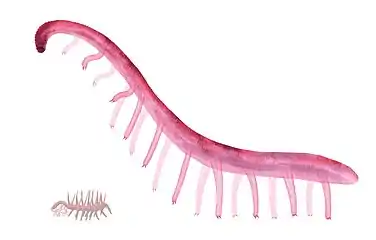 Paucipodia inermis (right) and Hallucigenia sparsa (bottom left) in scale.
Paucipodia inermis (right) and Hallucigenia sparsa (bottom left) in scale. Fossils of Xenusion, a lobopodian that may have grown up to 20 centimeters.
Fossils of Xenusion, a lobopodian that may have grown up to 20 centimeters.
Most lobopodians are only a few centimeters in length, while some genera may have grown up to over 20 centimeters.[6] Their bodies are annulated, although the annulation may be difficult to discern, due to their close spacing (~0.2 mm) and low relief on the fossil materials.[27] Body and appendages are circular in cross-section.[27]
Lobopod

The limbs of lobopodians, technically called lobopods[4] or lobopodous limbs,[12] are loosely conical in shape, tapering from the body to their clawed[27][15] or claw-lacking[6][28][29][13] tips. Usually the longest and most robust legs are at the middle of the trunk, with those nearer the anterior and posterior more spindly.[27] The claws, if present, are slightly curved, and their length is loosely proportional to the length of the leg to which they are attached.[27] In some genera, the lobopods may bear additional structures such as spines (e.g. Diania[29]), fleshy outgrowths (e.g. Onychodictyon), or tubercules (e.g. Jianshanopodia[6]). There's no signs of arthropodization (development of harden exoskeleton and segmental division on panarthropod appendages) in known members of lobopodians even for those belong to the arthropod stem-group (e.g. gilled lobopodians and siberiids), and the suspected case of arthropodization on the limbs of Diania[30] is considered to be a misinterpretation.[29][13]
Head

Heads are more or less bulbous,[4] and may bear a pair of pre-ocular, protocerebral[18] appendages – for examples primary antennae[28][26][18][13] or well-developed frontal appendages,[4][12][31][6] which are individualized from the trunk lobopods[18][32] (with the exception of Antennacanthopodia, which have two pairs of head appendages instead of one[28]). Mouthparts may consist of rows of teeth[27][16][31][6][33] or conical proboscis.[34][4] The eyes may be represented by one or be numerous[35] pairs of simple ocelli[4] as has been shown in Paucipodia,[27] Luolishania,[26] Miraluolishania,[35] Ovatiovermis,[36] Onychodictyon,[34] Hallucigenia,[16] and possibly Aysheaia as well.[34] However in gilled lobopodians like Kerygmachela, the eyes are relatively complex reflective patches.[37][38]
Trunk
The trunk is elongated and compose of numerous body segments (somites) each bore a pair of lobopod, but the segmental boundaries are not externally significant as those of arthropods. The trunk segments may bear other external structures such as turbecles (e.g. Hadranax,[5] Kerygmachela[12]), spine/plate-like sclerites (e.g. armoured lobopodians[4]) or flaps (e.g. gilled lobopodians[12][33]). Differentiation between trunk appendages barely occur, except in luolishaniids and hallucigenids, where numerous pairs of their anterior lobopods are significantly slender and/or setose contrast to their posterior counterparts.[4][36] The trunk may terminate with a pair of lobopod (e.g. Aysheaia, Hallucigenia)[16] or tail-like extension (e.g. Siberion, Jianshanopodia).[6][39]
Internal structures

The gut of lobopodians is often straight, undifferentiated,[40] and sometimes preserved in the fossil record in three dimensions. In some specimens the gut is found to be filled with sediment.[27] The gut consists of a central tube occupying the full length of the lobopodian's trunk,[6] which does not change much in width - at least not systematically. However in some groups, specifically the gilled lobopodians and siberiids, the guts were surrounded by pairs of serially repeated kidney-shaped gut diverticulae (digestive glands).[6][31][40] In some specimens, parts of the lobopodian gut can be preserved in three dimensions. This cannot result from phosphatisation, which is usually responsible for 3-D gut preservation,[41] for the phosphate content of the guts is under 1%; the contents comprise quartz and muscovite.[27] The gut of the representative Paucipodia is variable in width, being widest at the centre of the body. Its position in the body cavity is only loosely fixed, so flexibility is possible.

No much are known about the neural anatomy of lobopodians due to the spare and mostly ambiguous fossil evidence. Possible traces of brain and ventral nerve cord were found in Megadictyon and Paucipodia, respectively.[31][27] The first confirmed evidence of lobopodian neural structures comes from the gilled lobopodian Kerygmachela in a 2018 study — it presents a brain compose of only protocerebrum (the frontalmost cerebral ganglion of panarthropods) which were directly connect to the nerves of eyes and frontal appendages, suggest the protocerebral ancestry of the head of lobopodians as well as panarthropods.[37]
In some extant ecdysozoan such as priapulids and onychophorans, there is a layer of outermost circular muscles and a layer of innermost longitudinal muscles. The onychophorans also has a third intermediate layer of interwoven oblique muscles between the two others. But the musculature of a fossil lobopodian Tritonychus show the opposite pattern; it is the outermost muscles that are longitudinal and the innermost layer consisting of circular muscles.[42]
Categories
Based on external morphology, lobopdians may fall under different categories — for example the general worm-like taxa as "xenusiid" or "xenusian"; xenusiid with sclerite as "armoured lobopodians"; and taxa with both robust frontal appendages and lateral flaps as "gilled lobopodians". Some of them were originally defined under a taxonomic sense (e.g. class Xenusia), but neither any of them are generally accepted as monophyletic in further studies.[15][17][18]
Armoured lobopodians


Armoured lobopodians referred to xenusiid lobopodians which bore repeated sclerites such as spine or plates on their trunk (e.g. Hallucigenia, Microdictyon, Luolishania) or lobopods (e.g. Diania). In contrast, lobopodians without sclerites may be referred to as "unarmoured lobopodians".[28][13] Function of the sclerites were interpreted as protective armor and/or muscle attachment points.[43][4] In some cases, only the disarticulated sclerites of the animal were preserved, which represented as component of small shelly fossils (SSF).[10] Armoured lobopodians were suggest to be onychophoran-related and may even represent a clade in some previous studies,[43] but their phylogenetic positions in later studies are controversial. (see text)
Gilled lobopodians

Dinocaridids with lobopodian affinities (due to shared features like annulation & lobopods) are referred to as "Gilled lobopodians". These forms sport a pair of flaps on each trunk segment, but otherwise no signs of arthropodization, in contrast to more derived dinocaridids like the Radiodonta that have robust and sclerotized frontal appendages. Gilled lobopodians cover at least two genera: Pambdelurion and Kerygmachela.[13] Opabinia may also fall under this category in a broader sense,[17][44] although the presence of lobopods in this genus is not definitively proven.[45] Omnidens, a genus known only from a Pambdelurion-like mouth apparatus, may also be a gilled lobopodian.[33] The body flaps may have functioned as both swimming appendages and gills,[14] and are possibly homologous to the dorsal flaps of radiodonts and exopods of Euarthropoda.[12][44] Whether these genera were true lobopodians is still contested by some.[46] However, they are widely accepted as stem-group arthropods just basal to radiodonts.[17][15][16][18]
Siberion and similar taxa

Siberion, Megadictyon and Jianshanopodia may be grouped taxonomically as Siberiida or siberiids by some studies.[39] They are generally large (body length ranging between 7[39] and 22 centimeters[31]) xenusiid lobopodians with widen trunk, stout trunk lobopods without evidence of claws, and most notably a pair of robust frontal appendages.[17] With the exception of Siberion, they also have digestive glands like those of a gilled lobopodian and basal euarthropod.[17][40] Their anatomy represent transitional froms between typical xenusiids and gilled lobopodians,[39] eventually placing them under the basalmost position of arthropod stem-group.[6][31][17][18]
Paleoecology
Lobopodians may have occupied a wide range of ecological niches. Although most of them had undifferentiated appendages and straight gut, which would suggest a simple sediment-feeding lifestyle,[4] sophisticated digestive glands and large size of gilled lobopodians and siberiids would allow them to consume larger food items,[4][40] and their robust frontal appendages may even suggest a predatory lifestyle.[6][40] On the other hand, luolishaniids such as Luolishania and Ovatiovermis have elaborate feather-like lobopods that presumably formed 'baskets' for suspension or filter-feeding.[26][36] Lobopods with curved termial claws may have given some lobopodians the ability to climb on substrances.[4]
Not much is known about the physiology of lobopodians. There are evidence suggest that lobopodians moult just like other ecdysozoan taxa, but the outline and ornamentation of the harden sclerite did not vary during ontogeny.[11] The gill-like structures on the body flaps of gilled lobopodians and ramified extensions on the lobopods of Jianshanopodia may provide respiratory function (gills).[14][6] Pambdelurion may control the movement of their lobopods in a way similar to onychophorans.[47]
Distribution
During the Cambrian, lobopodians displayed a substantial degree of biodiversity. One species is known from each of the Ordovician and Silurian periods,[8][48] with a few more known from the Carboniferous (Mazon Creek) — this represents the paucity of exceptional lagerstatten in post-Cambrian deposits.
Phylogeny
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Neutralized phylogeny between lobopodians and other Ecdysozoan taxa.[15][17][16][44][18][36][19] Extant panarthropod taxa are in bold. Relationship between the total-group of extant panarthropod phyla is unresolve. |



The overall phylogenetic interpertation on lobopodians changed dramatically beyond decades.[4] The reassignments are not only based on new fossil evidence, but also new embryological, neuroanatomical, and genomic (e.g. gene expression, phylogenomics) informations observed from extant panarthropod taxa.[4][18][49]
Based on their apparently onychophoran-like morphology (e.g. annulated cuticle, lobopodous appendage with claws), lobopodians were traditionally thought to be present a group of paleozoic onychophorans.[4][50][51][52] This interpretation was flawed after the discovery of lobopodians with arthropod and tardigrade-like characters,[53] suggest the similarity between lobopodians and onychophorans represent deeper panarthropod ancestral trait (plesiomorphies) instead of onychophoran-exclusive characters (synamorphies).[19] For example, The British palaeontologist Graham Budd sees the Lobopodia as representing a basal grade from which the phyla Onychophora and Arthropoda arose, with Aysheaia comparable to the ancestral plan, and with forms like Kerygmachela and Pambdelurion representing a transition that, via dinocaridids to arthropods, would lead to an arthropod body plan.[43] Aysheaia's surface ornamentation, if homologous with palaeoscolecid sclerites, may represent a deeper link connecting it with cycloneuralian outgroups.[43] Many further studies follow and extend the idea, generally agreed that all three panarthropod phyla have lobopodians in their stem lineages.[4][15][16][18][36][19] Lobopodians are thus paraphyletic, and include the last common ancestor of arthropods, onychophorans and tardigrades.[4]
As stem-group arthropods
Compared to other panarthropod stem-groups, suggestion on the lobopodian members of arthropod stem-group is relatively consistent — siberiid like Megadictyon and Jianshanopodia occupied the basalmost position, gilled lobopodians Pambdelurion and Kerygmachela branch next, and finally lead to a clade compose of Opabinia, Radiodonta and Euarthropoda (crown-group arthropods).[15][17][16][44][18][36][19] Their positions within arthropod stem-group are indicated by numerous arthropod groundplans and intermediate forms (e.g. arthropod-like digestive glands, radiodont-like frontal appendages and dorso-ventral appendicular structures link to arthropod biramus appendages).[17][18] Lobopodian ancestry of arthropods also reinforced by genomic studies on extant taxa — gene expression support the homology between arthropod appendages and onychophoran lobopods, suggests that modern less-segmented arthropodized appendages evolved from annulated lobopodous limbs, with multipodomerous appendages of extinct basal euarthropods (e.g. fuxianhuiids) may represent an intermediate form.[32] On the other hand, primary antennae and frontal appendages of lobopodians and dinocaridids may be homologous to the labrum/hypostome complex of euarthropods, an idea support by their protocerebral origin[17][18][37] and developmental pattern of the labrum of extant arthropods.[32][18]
 Radiodonts are stem-group arthropods with gilled lobopodian-like body flaps, arthropodized frontal appendages and stalked compound eyes.
Radiodonts are stem-group arthropods with gilled lobopodian-like body flaps, arthropodized frontal appendages and stalked compound eyes.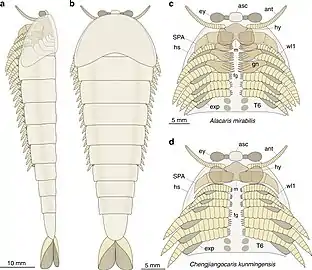 The multisegmented limbs of fuxianhuiid may represent intermediate form between lobopods and modern arthropod appendages.
The multisegmented limbs of fuxianhuiid may represent intermediate form between lobopods and modern arthropod appendages.
Diania, a genus of armoured lobopodian with stout and spiny legs, were originally thought to be associated within the arthropod stem-group based on its apparently arthropod-like (arthropodized) trunk appendages.[30] However, this interpretation is questionable as the data provided by the original description are not consistent with the suspected phylogenic relationships.[54][55] Further re-examination even revealed that the suspcted arthropodization on the legs of Diania was a misinterpretation — although the spine may have hardened, the remaining cuticle of Diania's legs were soft (not harden nor scleritzed), lacking any evidence of pivot joint and arthrodial membrane, suggest the legs are lobopods with only widely-spaced annulations.[29][13] Thus, the re-examination eventually reject the evidence of arthropodization (sclerotization, segmentation and articulation) on the appendages as well as the fundamental relationship between Diania and arthropods.[29][13]
As stem-group onychophorans

While Antennacanthopodia is widely accepted as a member of stem-group onychophoran,[15][16][18][36][19] position of other xenusiid genera that previously though to be onychophoran-related are controversial — in further studies, most of them were either suggest to be stem-group onychophorans[4][15][18] or basal panarthropods,[36][19] with a few species occasionally suggest to be stem-group tardigrades and/or stem-group panarthropods.[36][19] A study in 2014 suggest that Hallucigenia are stem-group onychophorans based on their claws, which have overlapped internal structures resemble to those of an extant onychophoran.[15] This interpretation was questioned by later studies, as the structures may present panarthropod plesiomorphy.[19]
As stem-group tardigrades
Lobopodian taxa of tardigrade stem-group is unclear.[4] Aysheaia[36][19] or Onychodictyon ferox[15][16] had been suggest to be a possible member. Although not widely accepted, there are even suggestions that Tardigrada itself representing the basalmost panarthropod or branch between the arthropod stem-group.[53]
As stem-group panarthropods
It is unclear that which lobopodians represent members of the panarthropod stem-group, which were branched just before the last common ancestor of extant panarthropod phyla. Aysheaia may have occupied this position based on its apparently basal morphology;[43][15][16] while other studies rather suggest luolishaniid and hallucigenid,[36][19] two lobopodian taxa which had been resolved as members of stem-group onychophorans as well.[4][15][18]
Described genera




As of 2018, over 20 lobopodian genera have been described.[13] The fossil materials being described as lobopodians Mureropodia apae and Aysheaia prolata are considered to be disarticulated frontal appendages of the radiodonts Caryosyntrips and Stanleycaris, respectively.[56][57][58] Miraluolishania was suggested to be synonym of Luolishania by some authors.[59][60] The enigmatic Facivermis was later revealed to be a highly specialized genus of luolishaniid lobopodians.[39][36][61]
- Antennacanthopodia
- Aysheaia
- Carbotubulus
- Cardiodictyon
- Collinsium
- Collinsovermis[62]
- Diania
- Facivermis
- Hadranax
- Hallucigenia
- Jianshanopodia
- Kerygmachela
- Lenisambulatrix
- Luolishania
- Megadictyon
- Microdictyon
- Miraluolishania (=Luolishania?)
- Onychodictyon
- Pambdelurion
- Paucipodia
- Siberion
- Thanahita
- Tritonychus
- Ovatiovermis
- Orstenotubulus
- Xenusion
References
- Haug, J.T.; Mayer, G.; Haug, C.; Briggs, D.E.G. (2012). "A Carboniferous non-Onychophoran Lobopodian reveals long-term survival of a Cambrian morphotype". Current Biology. 22 (18): 1673–1675. doi:10.1016/j.cub.2012.06.066. PMID 22885062.
- Snodgrass, R.E. (1938). "Evolution of the Annelida, Onychophora, and Arthropoda". Smithsonian Miscellaneous Collections. 97 (6): 1–159.
- Cavalier-Smith, T. (1998). "A revised six-kingdom system of life". Biological Reviews. 73 (3): 203–266. doi:10.1017/S0006323198005167. PMID 9809012.
- Ortega-Hernández, Javier (2015-10-05). "Lobopodians". Current Biology. 25 (19): R873–R875. doi:10.1016/j.cub.2015.07.028. ISSN 0960-9822. PMID 26439350.
- Budd, Graham; Peel, John (1998-12-01). "A new Xenusiid lobopod from the early Cambrian Sirius Passet fauna of North Greenland". Palaeontology. 41: 1201–1213.
- Jianni Liu; Degan Shu; Jian Han; Zhifei Zhang & Xingliang Zhang (2006). "A large xenusiid lobopod with complex appendages from the Lower Cambrian Chengjiang Lagerstätte" (PDF). Acta Palaeontol. Pol. 51 (2): 215–222. Retrieved 9 February 2011.
- van Roy, P.; Orr, P.J.; Botting, J.P.; Muir, L.A.; Vinther, J.; Lefebvre, B.; Hariri, K.E.; Briggs, D.E.G. (2010). "Ordovician faunas of Burgess Shale type". Nature. 465 (7295): 215–218. Bibcode:2010Natur.465..215V. doi:10.1038/nature09038. PMID 20463737. S2CID 4313285.
- von Bitter, P.H.; Purnell, M.A.; Tetreault, D.K.; Stott, C.A. (2007). "Eramosa Lagerstätte—Exceptionally preserved soft-bodied biotas with shallow-marine shelly and bioturbating organisms (Silurian, Ontario, Canada)". Geology. 35 (10): 879. Bibcode:2007Geo....35..879V. doi:10.1130/G23894A.1. S2CID 11561169.
- Haug, J.T.; Mayer, G.; Haug, C.; Briggs, D.E.G. (2012). "A Carboniferous non-Onychophoran Lobopodian reveals long-term survival of a Cambrian morphotype". Current Biology. 22 (18): 1673–1675. doi:10.1016/j.cub.2012.06.066. PMID 22885062.
- Caron, J.-B.; Smith, M.R.; Harvey, T.H.P. (2013). "Beyond the Burgess Shale: Cambrian microfossils track the rise and fall of hallucigeniid lobopodians". Proceedings of the Royal Society B: Biological Sciences. 280 (1767): 20131613. doi:10.1098/rspb.2013.1613. PMC 3735267. PMID 23902914.
- Topper, T.P.; Skovsted, C.B.; Peel, J.S.; Harper, D.A.T. (2013). "Moulting in the lobopodian Onychodictyonfrom the lower Cambrian of Greenland". Lethaia: n/a. doi:10.1111/let.12026.
- Budd, Graham E. (1998). "The morphology and phylogenetic significance of Kerygmachela kierkegaardi Budd (Buen Formation, Lower Cambrian, N Greenland)". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 89 (4): 249–290. doi:10.1017/S0263593300002418. ISSN 1473-7116.
- Ou, Qiang; Mayer, Georg (2018-12-01). "A Cambrian unarmoured lobopodian, †Lenisambulatrix humboldti gen. et sp. nov., compared with new material of †Diania cactiformis". Scientific Reports. 8 (1): 13667. Bibcode:2018NatSR...813667O. doi:10.1038/s41598-018-31499-y. PMC 6147921. PMID 30237414.
- Budd, Graham (August 1993). "A Cambrian gilled lobopod from Greenland". Nature. 364 (6439): 709–711. Bibcode:1993Natur.364..709B. doi:10.1038/364709a0. ISSN 1476-4687. S2CID 4341971.
- Smith, Martin R.; Ortega-Hernández, Javier (2014). "Hallucigenia's onychophoran-like claws and the case for Tactopoda" (PDF). Nature. 514 (7522): 363–366. Bibcode:2014Natur.514..363S. doi:10.1038/nature13576. PMID 25132546. S2CID 205239797.
- Smith, Martin R.; Caron, Jean-Bernard (July 2015). "Hallucigenia's head and the pharyngeal armature of early ecdysozoans". Nature. 523 (7558): 75–78. Bibcode:2015Natur.523...75S. doi:10.1038/nature14573. ISSN 0028-0836. PMID 26106857. S2CID 205244325.
- Ortega-Hernández, Javier (December 2014). "Making sense of 'lower' and 'upper' stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848". Biological Reviews of the Cambridge Philosophical Society. 91 (1): 255–273. doi:10.1111/brv.12168. ISSN 1469-185X. PMID 25528950. S2CID 7751936.
- Ortega-Hernández, Javier; Janssen, Ralf; Budd, Graham E. (2017-05-01). "Origin and evolution of the panarthropod head – A palaeobiological and developmental perspective". Arthropod Structure & Development. Evolution of Segmentation. 46 (3): 354–379. doi:10.1016/j.asd.2016.10.011. ISSN 1467-8039. PMID 27989966.
- Siveter, Derek J.; Briggs, Derek E. G.; Siveter, David J.; Sutton, Mark D.; Legg, David (2018). "A three-dimensionally preserved lobopodian from the Herefordshire (Silurian) Lagerstätte, UK". Royal Society Open Science. 5 (8): 172101. doi:10.1098/rsos.172101. PMC 6124121. PMID 30224988.
- Dzik, Jerzy; Krumbiegel, Günter (1989). "The oldest 'onychophoran' Xenusion: a link connecting phyla?". Lethaia. 22 (2): 169–181. doi:10.1111/j.1502-3931.1989.tb01679.x. ISSN 1502-3931.
- Smith, Frank W.; Goldstein, Bob (2017-05-01). "Segmentation in Tardigrada and diversification of segmental patterns in Panarthropoda". Arthropod Structure & Development. Evolution of Segmentation. 46 (3): 328–340. doi:10.1016/j.asd.2016.10.005. ISSN 1467-8039. PMID 27725256.
- Pentastomida - Sociedad Entomológica Aragonesa
- Treatise on Zoology - Anatomy, Taxonomy, Biology. The Crustacea, Volum 5
- Fortey, Richard A.; Thomas, Richard H. (1997-12-31). Arthropod Relationships. Springer Science & Business Media. ISBN 978-0-412-75420-3.
- Gould, S.J. (1989). Wonderful Life: The Burgess Shale and the Nature of History. W.W. Norton & Company. Bibcode:1989wlbs.book.....G.
- Ma, Xiaoya; Hou, Xianguang; Bergström, Jan (2009). "Morphology of Luolishania longicruris (Lower Cambrian, Chengjiang Lagerstätte, SW China) and the phylogenetic relationships within lobopodians". Arthropod Structure & Development. 38 (4): 271–291. doi:10.1016/j.asd.2009.03.001. ISSN 1467-8039. PMID 19293001.
- Hou, Xian-Guang; Ma, Xiao-Ya; Zhao, Jie; Bergström, Jan (2004). "The lobopodian Paucipodia inermis from the Lower Cambrian Chengjiang fauna, Yunnan, China". Lethaia. 37 (3): 235–244. doi:10.1080/00241160410006555.
- Ou, Qiang; Liu, Jianni; SHU, DEGAN; Han, Jian; Zhang, Zhifei; Wan, Xiaoqiao; Lei, Qianping (2011-05-01). "A Rare Onychophoran-Like Lobopodian from the Lower Cambrian Chengjiang Lagerstätte, Southwestern China, and its Phylogenetic Implications". Journal of Paleontology. 85 (3): 587–594. doi:10.1666/09-147R2.1. JSTOR 23020193. S2CID 53056128.
- Ma, Xiaoya; Edgecombe, Gregory; Legg, David; Hou, Xianguang (2013-05-08). "The morphology and phylogenetic position of the Cambrian lobopodian Diania cactiformis". Journal of Systematic Palaeontology. 12 (4): 445–457. doi:10.1080/14772019.2013.770418. S2CID 220463025.
- Liu, Jianni; Steiner, Michael; Dunlop, Jason A.; Keupp, Helmut; Shu, Degan; Ou, Qiang; Han, Jian; Zhang, Zhifei; Zhang, Xingliang (February 2011). "An armoured Cambrian lobopodian from China with arthropod-like appendages". Nature. 470 (7335): 526–530. Bibcode:2011Natur.470..526L. doi:10.1038/nature09704. ISSN 1476-4687. PMID 21350485. S2CID 4324509.
- Liu, J.; Shu, D.; Han, J.; Zhang, Z.; Zhang, X. (2007). "Morpho-anatomy of the lobopod Magadictyon cf. Haikouensis from the Early Cambrian Chengjiang Lagerstätte, South China". Acta Zoologica. 88 (4): 279–288. doi:10.1111/j.1463-6395.2007.00281.x.
- Jockusch, Elizabeth L. (2017-09-01). "Developmental and Evolutionary Perspectives on the Origin and Diversification of Arthropod Appendages". Integrative and Comparative Biology. 57 (3): 533–545. doi:10.1093/icb/icx063. ISSN 1540-7063. PMID 28957524.
- Vinther, Jakob; Porras, Luis; Young, Fletcher; Budd, Graham; Edgecombe, Gregory (2016-09-01). "The mouth apparatus of the Cambrian gilled lobopodian Pambdelurion whittingtoni". Palaeontology. 59 (6): 841–849. doi:10.1111/pala.12256.
- Ou, Qiang; Shu, Degan; Mayer, Georg (2012-12-11). "Cambrian lobopodians and extant onychophorans provide new insights into early cephalization in Panarthropoda". Nature Communications. 3 (1): 1261. Bibcode:2012NatCo...3.1261O. doi:10.1038/ncomms2272. ISSN 2041-1723. PMC 3535342. PMID 23232391.
- Schoenemann, Brigitte; Liu, Jian-Ni; Shu, De-Gan; Han, Jian; Zhang, Zhi-Fei (2009). "A miniscule optimized visual system in the Lower Cambrian". Lethaia. 42 (3): 265–273. doi:10.1111/j.1502-3931.2008.00138.x. ISSN 1502-3931.
- Caron, Jean-Bernard; Aria, Cédric (2017-01-31). "Cambrian suspension-feeding lobopodians and the early radiation of panarthropods". BMC Evolutionary Biology. 17 (1): 29. doi:10.1186/s12862-016-0858-y. ISSN 1471-2148. PMC 5282736. PMID 28137244.
- Park, Tae-Yoon S.; Kihm, Ji-Hoon; Woo, Jusun; Park, Changkun; Lee, Won Young; Smith, M. Paul; Harper, David A. T.; Young, Fletcher; Nielsen, Arne T. (2018-03-09). "Brain and eyes of Kerygmachela reveal protocerebral ancestry of the panarthropod head". Nature Communications. 9 (1): 1019. Bibcode:2018NatCo...9.1019P. doi:10.1038/s41467-018-03464-w. ISSN 2041-1723. PMC 5844904. PMID 29523785.
- Fleming, James F.; Kristensen, Reinhardt Møbjerg; Sørensen, Martin Vinther; Park, Tae-Yoon S.; Arakawa, Kazuharu; Blaxter, Mark; Rebecchi, Lorena; Guidetti, Roberto; Williams, Tom A.; Roberts, Nicholas W.; Vinther, Jakob (2018-12-05). "Molecular palaeontology illuminates the evolution of ecdysozoan vision". Proceedings of the Royal Society B: Biological Sciences. 285 (1892): 20182180. doi:10.1098/rspb.2018.2180. ISSN 0962-8452. PMC 6283943. PMID 30518575.
- Dzik, Jerzy (1 July 2011). "The xenusian-to-anomalocaridid transition within the lobopodians" (PDF). Bollettino della Societa Paleontologica Italiana. 50: 65–74.
- Vannier, Jean; Liu, Jianni; Lerosey-Aubril, Rudy; Vinther, Jakob; Daley, Allison C. (2014-05-02). "Sophisticated digestive systems in early arthropods". Nature Communications. 5 (1): 3641. Bibcode:2014NatCo...5.3641V. doi:10.1038/ncomms4641. ISSN 2041-1723. PMID 24785191.
- Butterfield, N. J. (2002). "Leanchoilia guts and the interpretation of three-dimensional structures in Burgess Shale-type fossils". Paleobiology. 28: 155–171. doi:10.1666/0094-8373(2002)028<0155:LGATIO>2.0.CO;2. ISSN 0094-8373.
- Onychophoran-like musculature in a phosphatized Cambrian lobopodian
- Budd, G. E. (2001). "Why are arthropods segmented?". Evolution and Development. 3 (5): 332–42. doi:10.1046/j.1525-142X.2001.01041.x. PMID 11710765. S2CID 37935884.
- Van Roy, Peter; Daley, Allison C.; Briggs, Derek E. G. (2015). "Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps". Nature. 522 (7554): 77–80. Bibcode:2015Natur.522...77V. doi:10.1038/nature14256. ISSN 1476-4687. PMID 25762145. S2CID 205242881.
- Budd, Graham E.; Daley, Allison C. (January 2012). "The lobes and lobopods of Opabinia regalis from the middle Cambrian Burgess Shale: The lobes of Opabinia". Lethaia. 45 (1): 83–95. doi:10.1111/j.1502-3931.2011.00264.x.
- Chen, J.-y; Ramskold, L.; Zhou, G.-q (1994). "Evidence for Monophyly and Arthropod Affinity of Cambrian Giant Predators". Science. 264 (5163): 1304–1308. Bibcode:1994Sci...264.1304C. doi:10.1126/science.264.5163.1304. ISSN 0036-8075. PMID 17780848. S2CID 1913482.
- Young, Fletcher J.; Vinther, Jakob (2017). "Onychophoran-like myoanatomy of the Cambrian gilled lobopodian Pambdelurion whittingtoni" (PDF). Palaeontology. 60 (1): 27–54. doi:10.1111/pala.12269. ISSN 1475-4983.
- Whittle, R. J.; Gabbott, S. E.; Aldridge, R. J.; Theron, J. (2009). "An Ordovician Lobopodian from the Soom Shale Lagerstätte, South Africa". Palaeontology. 52 (3): 561–567. doi:10.1111/j.1475-4983.2009.00860.x.
- Edgecombe, Gregory D. (2009-06-01). "Palaeontological and Molecular Evidence Linking Arthropods, Onychophorans, and other Ecdysozoa". Evolution: Education and Outreach. 2 (2): 178–190. doi:10.1007/s12052-009-0118-3. ISSN 1936-6434.
- Ramsköld, L.; Xianguang, Hou (1991). "New early Cambrian animal and onychophoran affinities of enigmatic metazoans". Nature. 351 (6323): 225–228. Bibcode:1991Natur.351..225R. doi:10.1038/351225a0. ISSN 1476-4687. S2CID 4309565.
- Bergström, J.; Hou, Xian-Guang (2001-12-01). "Cambrian Onychophora or Xenusians". Zoologischer Anzeiger - ZOOL ANZ. 240 (3–4): 237–245. doi:10.1078/0044-5231-00031.
- Robison, R. A. (1985). "Affinities of Aysheaia (Onychophora), with Description of a New Cambrian Species". Journal of Paleontology. 59 (1): 226–235. ISSN 0022-3360. JSTOR 1304837.
- Budd, Graham E. (2001-01-01). "Tardigrades as 'Stem-Group Arthropods': The Evidence from the Cambrian Fauna". Zoologischer Anzeiger - A Journal of Comparative Zoology. 240 (3): 265–279. doi:10.1078/0044-5231-00034. ISSN 0044-5231.
- Mounce, Ross C. P.; Wills, Matthew A. (August 2011). "Phylogenetic position of Diania challenged". Nature. 476 (7359): E1, discussion E3–4. Bibcode:2011Natur.476E...1M. doi:10.1038/nature10266. ISSN 1476-4687. PMID 21833044. S2CID 4417903.
- Legg, David; Ma, Xiaoya; Wolfe, Joanna; Ortega-Hernández, Javier; Edgecombe, Gregory; Sutton, Mark (2011-08-11). "Lobopodian phylogeny reanalysed". Nature. 476 (7359): E2–3, discussion E3. Bibcode:2011Natur.476Q...1L. doi:10.1038/nature10267. PMID 21833046. S2CID 4310063.
- Pates, Stephen; Daley, Allison C. (2017). "Caryosyntrips: a radiodontan from the Cambrian of Spain, USA and Canada". Papers in Palaeontology. 3 (3): 461–470. doi:10.1002/spp2.1084. ISSN 2056-2802.
- "Aysheaia prolata from the Utah Wheeler Formation (Drumian, Cambrian) is a frontal appendage of the radiodontan Stanleycaris - Acta Palaeontologica Polonica". www.app.pan.pl. Retrieved 2020-01-08.
- "Reply to Comment on "Aysheaia prolata from the Utah Wheeler Formation (Drumian, Cambrian) is a frontal appendage of the radiodontan Stanleycaris" with the formal description of Stanleycaris - Acta Palaeontologica Polonica". www.app.pan.pl. Retrieved 2020-01-08.
- Ma, Xiaoya; Hou, Xianguang; Bergström, Jan (2009-07-01). "Morphology of Luolishania longicruris (Lower Cambrian, Chengjiang Lagerstätte, SW China) and the phylogenetic relationships within lobopodians". Arthropod Structure & Development. 38 (4): 271–291. doi:10.1016/j.asd.2009.03.001. ISSN 1467-8039. PMID 19293001.
- Liu, Jianni; Shu, DeGan; Han, Jian; Zhang, Zhifei (2008-05-01). "Comparative study of Cambrian lobopods Miraluolishania and Luolishania". Chinese Science Bulletin - CHIN SCI BULL. 53 (1): 87–93. Bibcode:2008ChSBu..53...87L. doi:10.1007/s11434-007-0428-1. S2CID 128619311.
- Howard, Richard J.; Hou, Xianguang; Edgecombe, Gregory D.; Salge, Tobias; Shi, Xiaomei; Ma, Xiaoya (2020-02-27). "A Tube-Dwelling Early Cambrian Lobopodian". Current Biology. 0 (8): 1529–1536.e2. doi:10.1016/j.cub.2020.01.075. ISSN 0960-9822. PMID 32109391.
- Caron, Jean-Bernard; Aria, Cédric (2020). "The Collins' monster, a spinous suspension-feeding lobopodian from the Cambrian Burgess Shale of British Columbia". Palaeontology. n/a (n/a). doi:10.1111/pala.12499. ISSN 1475-4983.