Sparassodonta
Sparassodonta (from Greek σπαράσσειν [sparassein], to tear, rend; and ὀδούς, gen. ὀδόντος [odous, odontos], tooth) is an extinct order of carnivorous metatherian mammals native to South America. They were once considered to be true marsupials, but are now thought to be either a sister taxon to them,[1][2][3][4] or considerably distantly related, part of a separate clade of Gondwanan metatherians.[5] A number of these mammalian predators closely resemble placental predators that evolved separately on other continents, and are cited frequently as examples of convergent evolution. They were first described by Florentino Ameghino, from fossils found in the Santa Cruz beds of Patagonia. Sparassodonts were present throughout South America's long period of "splendid isolation" during the Cenozoic; during this time, they shared the niches for large warm-blooded predators with the flightless terror birds. Previously, it was thought that these mammals died out in the face of competition from "more competitive" placental carnivorans during the Pliocene Great American Interchange, but more recent research has showed that sparassodonts died out long before eutherian carnivores arrived in South America (aside from procyonids, which sparassodonts probably did not directly compete with).[6][7][8] Sparassodonts have been referred to as borhyaenoids by some authors,[9][10][11] but currently the term Borhyaenoidea refers to a restricted subgroup of sparassodonts comprising borhyaenids and their close relatives.[12][13]
| Sparassodonta | |
|---|---|
 | |
| Lycopsis | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Clade: | Metatheria |
| Clade: | Marsupialiformes |
| Order: | †Sparassodonta Ameghino, 1894 |
| Families | |
|
†Hathliacynidae | |
Features
Although members of the order Sparassodonta showed many similarities with placental carnivores, they were not closely related and are a very good example of convergent evolution. For example, sparassodonts' molars were very similar to the sharp teeth of placental carnivores. The canines were also lengthened, and in some cases resemble those of saber-toothed cats. Sparassodonts spanned a wide range of body sizes, from 2.2 pound (1 kg) weasel or civet-like forms to Thylacosmilus, which was the size of a leopard.[7][14] Along with the Australian thylacoleonids, sparassodonts include some of the largest metatherian carnivores.[7]
Sparassodonta is characterized by dental synapomorphies that distinguish the group from other closely related mammals. Unequivocal traits uniting the earliest Sparassodonts include:[12][15]
- a snout that forms a pronounced bulge around the canine teeth when viewed from above
- a ridge on the upper molar (preparacrista) oriented anterobuccally (towards the cheek) with respect to the long axis of the tooth.
- a pronounced keel near the base of the front of the paraconid
- ridges on lower molars (postprotocristid-metacristid) parallel or oblique with respect to lower jaw axis.
- a very tall protoconid (>90% tooth length, secondarily lost in Hondadelphys and Stylocynus) that bulges to the side and is wider at its midpoint than its base
- talonid (crushing end) of lower molar narrow in relation to trigonid (shearing end).
The dental formula of most sparassodonts is 4.1.3.43.1.3.4, with the exception of borhyaenids, which have three upper incisors, proborhyaenids (except for Callistoe), which have only two lower incisors, and thylacosmilids, which have only two lower incisors, at least two upper incisors, and only two upper and lower premolars on each side of the jaw.[12][16] Some specimens of Borhyaena and Arctodictis are also missing the last upper molar, showing that the presence of this tooth was variable in these species.[17] Sparassodonts had a typical metatherian mode of dental replacement, replacing only the upper and lower third premolar throughout their lifetime.[18] In thylacosmilids, the deciduous upper premolar was never replaced.[19]
Sparassodonts have highly reduced epipubic bones,[17] to the point that early analysis could not even find evidence for them.[20] This is a characteristic shared with the Australian thylacine, and historically argued as a synapomorphy,[17] though nowadays it is considered to have developed independently for poorly understood reasons. Like with thylacines, it is very likely that they possessed long cartilaginous elements instead.[10]
Evolutionary history
Sparassodonts have been suggested to be related to a variety of other groups of metatherians. Florentino Ameghino, who first described fossils of the group, thought that sparassodonts were closely related to creodonts and were a transitional group between metatherians and carnivorous placentals (including modern carnivorans).[21][12] Contemporary authors in the late 19th and early 20th century rejected this hypothesis and considered sparassodonts to be closely related to Australian thylacines and dasyurids.[22][23][24][25] The most popular hypothesis for much of the 20th century was that sparassodonts were closely related to opossums.[26][9] In 1990, Marshall et al. (1990) considered the Cretaceous stagodontids to be members of Sparassodonta, but this was criticized by later authors.[13] Marshall and Kielan-Jaworowska (1992) considered sparassodonts to be closely related to deltatheroidans, but this was also criticized.[27] Most of these hypotheses were based on similar adaptations for carnivorous diets in sparassodonts, opossums, dasyuromorphians, stagodonts, and deltatheroidans, which are highly prone to convergent evolution within mammals.[28] Szalay (1994) considered sparassodonts to be closely related to paucituberculatans based on features of the ankle.[29] In recent years there has been a growing consensus that sparassodonts are positioned just outside of crown-group Marsupialia, in a broader clade (Pucadelphyida) including pucadelphyids as well as sparassodonts.[12][15][30]
Several other metatherian taxa have been suggested to be sparassodonts or closely related to sparassodonts. The australian Murgon taxa Archaeonothos has been noted as being similar to sparassodonts, but currently its relationships are not fully concluded.[31] Carneiro (2018) recovered the genus Varalphadon from the Late Cretaceous of North America as a basal member of Sparassodonta.[32] However, this interpretation of Varalphadon as a sparassodont has not been supported by later phylogenetic analyses, and most of the purported synapomorphies between Varalphadon and sparassodonts are not actually present in Varalphadon[15] or have been suggested to be due to convergent evolution.[30] Sparassodonts are currently considered to be endemic to South America, and have not even been found in nearby continents like Antarctica (which otherwise shares many groups of mammals with South America such as litopterns, astrapotheres, microbiotheres, and polydolopids).[33][30]
The early history of the Sparassodonta is poorly known, as most Paleocene and Eocene members of this group are only known from isolated teeth and fragmentary jaws. However, one species, the middle Eocene Callistoe vincei, is known from a nearly complete, articulated skeleton.[34] As Callistoe belongs to one of the most specialized groups of sparassodonts, this indicates that the other major groups (e.g. borhyaenids, hathliacynids, etc.) must have also arisen by this time. Originally, the early Paleocene metatherian Mayulestes was considered to be the earliest known member of the Sparassodonta, but phylogenetic analyses suggest that this species represents an independent radiation of carnivorous metatherians more closely related to Pucadelphys;[12] however, recent studies show that these taxa were closely related to borhyaenids.[35] As of this writing, the earliest known true sparassodonts are either Allqokirus australis, a species from the same site as Mayulestes that may turn out to not be a sparassodont, and an isolated astragalus from the earliest Paleocene site of Punta Peligro, Argentina.[36]
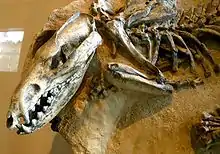
Sparassodonts can be divided into six major groups; basal sparassodonts (?earliest Paleocene-late Miocene), species that cannot be easily assigned to any of the other sparassodont groups and whose teeth often exhibit adaptations for omnivory; hathliacynids (late Oligocene-early Pliocene/late Pliocene), which range from a marten to a thylacine in size, and have long, fox-like snouts and teeth strongly adapted for carnivory; basal borhyaenoids (middle Eocene-late Miocene), borhyaenoids that cannot be easily classified into the families Borhyaenidae, Thylacosmilidae, or Proborhyaenidae and vary in size and shape; borhyaenids (early-late Miocene), the sparassodont group most specialized for running, but not as much as living carnivorans or even thylacines; proborhyaenids (middle Eocene-late Oligocene), robust, wolverine-like forms with ever-growing upper and lower canines; and thylacosmilids (early Miocene-late Pliocene), another terrestrially specialized group with ever-growing saber-like upper canines.[12]
After the middle Miocene, sparassodonts began to slowly decline in diversity. Basal borhyaenoids are last known from the early late Miocene (Pseudolycopsis cabrerai and Lycopsis viverensis), and after this time were at least partially replaced by large-bodied basal sparassodonts such as Stylocynus. It has been suggested that this shift in dominance was due to the more omnivorous habits of basal sparassodonts, which may have been better able to exploit the more seasonal climates of South America during the late Neogene.[37] Borhyaenids are last known from the latest Miocene, though only fragmentary remains of this group are known from this period.[6] Later remains assigned to this group have since been reidentified as thylacosmilids or procyonids. By the Pliocene, only two families of sparassodonts remained in South America, the Hathliacynidae and the Thylacosmilidae. Pliocene hathliacynid remains are rare, and it is possible that these animals may have competed with the large carnivorous didelphids such as Lutreolina that appeared around this time.[38] Hathliacynids are last definitively known from the early Pliocene, though their remains are rare. The thylacosmilids, on the other hand, were more successful and abundant, being some of the only large mammalian carnivores in South America during the Pliocene, before dying out during a faunal turnover in the middle of the epoch (the youngest specimens of thylacosmilids are ~3.3 Ma).[7] It is still not certain why Sparassodonta declined in diversity and became extinct during the late Cenozoic, but it appears as though competition from eutherian carnivorans was not a factor, as the placental analogues of sparassodonts (dogs, weasels, and saber-toothed cats) did not enter South America until the middle Pleistocene, several million years after their sparassodont counterparts became extinct.[6][12][39] Sparassodonts did coexist with Cyonasua-group procyonids during the late Miocene and Pliocene, but Cyonasua-group procyonids appear to have been primarily omnivorous and filled ecological niches that sparassodonts never occupied, which may be one reason that these animals were able to colonize South America despite the diverse predator guild in the late Miocene.[8] The overall decline in sparassodont diversity from the late Miocene to the end of the Pliocene may be linked to the cooling climates following the middle Miocene climatic optimum and the onset of the Pleistocene ice ages.[40]
Paleobiology
Sparassodonts were carnivorous, and with the exception of some basal members of all members of this group were hypercarnivorous (having diets composed of more than 70% meat).[7][41] Only Hondadelphys and Stylocynus appear to have exhibited adaptations for omnivory, and even then Stylocynus may have had a more mesocarnivorous diet similar to canids than an omnivorous one.[8] Medium-to-large caviomorph rodents and rodent-like mammals (e.g., small notoungulates) appear to have been common prey items of sparassodonts. The subadult holotype of Lycopsis longirostrus preserves remains of the dinomyid Scleromys colombianus as fossilized gut contents.[42] Sparassodont coprolites from the Santa Cruz Formation preserve the bones of chinchillid and octodontoid rodents inside them.[43] Bite marks from medium-sized sparassodonts have been found on the small notoungulate Paedotherium.[44] Stable isotope data from the early late Miocene Lycopsis viverensis and Thylacosmilus atrox suggests that these species fed on C3 grazers in open habitats, likely notoungulates.[45]
Bite marks likely pertaining to hathliacynid sparassodonts have been found on the remains of penguins and flightless marine ducks in ancient seabird nesting colonies, suggesting that sparassodonts raided seabird colonies for eggs, carrion, and other prey like many predatory mammals do today.[46][47]
Borhyaenid and proborhyaenid sparassodonts have been interpreted as being capable of crushing bones similar to modern hyenas, wolverines, or the Tasmanian devil (Sarcophilus harrisii) based on their deep jaws, bulbous premolars with deep roots and pronounced wear at their tips, extensive fused or interlocking mandibular symphyses, large masseteric fossae, microfractures in their tooth enamel, and high estimated bite forces.[48][49][50] Australohyaena antiquua shows particularly pronounced adaptations for bone-cracking, with a very deep jaw and strongly arched nasals similar to what is seen in modern hyaenids.[51]
Based on studies of the postcranial skeleton, it appears as though most sparassodonts were scansorial (adapted for climbing), although terrestrial adaptations evolved in Lycopsis longirostrus, borhyaenids, proborhyaenids, and thylacosmilids.[52][53][54] Most sparassodonts were plantigrade, Borhyaena has been suggested to have been digitigrade[55] but this has been questioned.[12] The one exception was Thylacosmilus, which has been interpreted as having a digitigrade forefoot and a semiplantigrade hindfoot,[55][56] this has been supported by fossil tracks.[57]
One unusual aspect of sparassodont paleoecology is that at most fossil localities their remains are nearly ten times rarer than would be expected based on comparisons with carnivorous mammals at fossil sites in other parts of the world.[58][59][60][61] The exact reasons for this are not clear, though this appears to be a broader pattern applicable to other groups of Cenozoic South American terrestrial carnivores (i.e., terror birds).[61]
Little is known of the behavior and biology of sparassodonts outside of general locomotor and dietary habits. Argot (2004) proposed that Thylacosmilus atrox may have exhibited protracted parental care after weaning of the offspring, given that saber teeth in general have been suggested to require long juvenile periods in order for the young to gain the skill necessary to use them effectively.[11] However, this has not been tested further. Sparassodonts have relatively large and complex brains for metatherians, comparable to those of some Australian marsupials like Australian possums,[62] though the body masses used to produce these estimates of relative brain size are low compared later studies suggesting these values could be overestimated.[63] Sparassodonts appear to have had very little binocular vision, with borhyaenids having the greatest degree of depth perception (but still lower than modern carnivorans) and the eyes of Thylacosmilus facing almost completely to the sides.[64] However, later studies have found that Thylacosmilus likely held its head in a downward-facing position, which would have allowed for more binocular vision than previously thought.[65] Wounds have been documented on the face of specimens of Borhyaena tuberata and Sipalocyon gracilis, potentially suggesting aggressive habits similar to the modern Tasmanian devil (Sarcophilus harrisii).[23] Several specimens of hathliacynids (Sipalocyon and Cladosictis) show a pathological disorder characterized by the presence of growths on the surface of the mandible, which in the most extreme cases can result in the loss of several teeth due to bony pathological growths.[17] The exact cause of this condition (i.e., infection, virus, parasite) and why it seems to only occur in small sparassodonts is unknown, though this condition has also been documented in microbiotherians.[66]
Classification
The following cladogram of sparassodont interrelationships is after Engelman et al., 2020.[67] Not all studies agree on the sister group relationship between Thylacosmilidae and Borhyaenidae recovered here, with other studies finding thylacosmilids to be more closely related to proborhyaenids.[34] The relationships among hathliacynids are also relatively unstable.[68]
| Sparassodonta |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Within Metatheria, a 2016 phylogenetic analysis group found that borhyaenids form a clade with the Asian "Gurlin Tsav skull" as well as other South American taxa. The same phylogeny found that marsupials group among various North American Cretaceous species. The phylogenetic tree is reproduced below.[69]
| Marsupialiformes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Taxonomy
The taxonomic classification below follows the latest review of the group, that of Prevosti and Forasiepi (2018),[70] with additions from more recent studies.[12] Although Mayulestes was originally described as a sparassodont, later phylogenetic analyses have shown that it most likely does not belong to this group;[12][71] however more recent studies show it to be closely related to sparassodonts.[35] Similarly, while basal borhyaenoids such as Lycopsis and Prothylacynus were once thought to belong to a distinct family (Prothylacynidae), phylogenetic analyses have found that these animals do not represent a monophyletic group.[12][34][72][73] The exact age of most Eocene species of sparassodonts is uncertain, given the lack of precise stratigraphic information associated with most specimens and the recent division of the Casamayoran SALMA into the Vacan and Barrancan SALMAs.
- Order Sparassodonta
- Genus Allqokirus[15]
- Genus Mayulestes[15]
- Genus Argyrolestes
- Genus Nemolestes
- Genus Patene
- Patene campbelli (late ?Eocene, Santa Rosa local fauna)
- Patene coloradensis (middle Eocene)[30]
- Patene coluapiensis (middle Eocene, Barrancan SALMA)
- Patene simpsoni (late Paleocene/early Eocene, Itaboraian SALMA)
- Genus Procladosictis
- Family Hondadelphyidae
- Genus Hondadelphys
- Genus Stylocynus
- Stylocynus paranensis (latest Miocene, Huayquerian SALMA)
- Family Hathliacynidae
- Genus Acyon
- Acyon ?herrerae (early Miocene, Colhuehuapian SALMA)
- Acyon myctoderos (late middle Miocene, Laventan SALMA)
- Acyon tricuspidatus (late early Miocene, Santacrucian SALMA)
- Genus Australogale
- Genus Borhyaenidium
- Borhyaenidium altiplanicus (latest Miocene, Huayquerian SALMA)
- Borhyaenidium riggsi (unknown, either early or late Pliocene, Montehermosan or Chapadmalalan SALMA)
- Borhyaenidium musteloides (latest Miocene, Huayquerian SALMA)
- Genus Chasicostylus
- Genus Cladosictis
- Cladosictis centralis (early Miocene, Colhuehuapian SALMA)
- Cladosictis patagonica (late early to earliest middle Miocene, Santacrucian-Friasian SALMA)
- Genus Notictis
- Notictis ortizi (latest Miocene, Huayquerian SALMA)
- Genus Notocynus
- Notocynus hermosicus (early Pliocene, Montehermosan SALMA)
- Genus Notogale
- Genus Pseudonotictis
- Pseudonotictis chubutensis (early middle Miocene, Colloncuran SALMA)
- Pseudonotictis pusillus (late early Miocene, Santacrucian SALMA)
- Genus Perathereutes
- Perathereutes pungens (late early Miocene, Santacrucian SALMA)
- Genus Sallacyon
- Genus Sipalocyon
- Sipalocyon externus (early Miocene, Colhuehuapian SALMA)
- Sipalocyon gracilis (late early Miocene, Santacrucian SALMA)
- Sipalocyon "obusta" (late early Miocene, Santacrucian SALMA)
- Genus Acyon
- Superfamily Borhyaenoidea
- Genus Angelocabrerus
- Angelocabrerus daptes (middle Eocene)
- Genus Chlorocyon
- Genus Dukecynus
- Genus Fredszalaya[16]
- Genus Lycopsis
- Genus Pharsophorus
- Genus Plesiofelis
- Genus Prothylacynus
- Prothylacynus patagonicus (late early to earliest middle Miocene, Santacrucian-Friasian SALMA)
- Genus Pseudolycopsis
- Genus Pseudothylacynus
- Pseudothylacynus rectus (early Miocene, Colhuehuapian SALMA)
- Family Borhyaenidae
- Genus Acrocyon
- Acrocyon riggsi (early Miocene, Colhuehuapian SALMA)
- Acrocyon sectorius (late early Miocene, Santacrucian SALMA)
- Genus Arctodictis
- Arctodictis munizi (late early Miocene, Santacrucian SALMA)
- Arctodictis sinclairi (early Miocene, Colhuehuapian SALMA)
- Genus Australohyaena
- Genus Borhyaena
- Borhyaena macrodonta (early Miocene, Colhuehuapian SALMA)
- Borhyaena tuberata (late early Miocene, Santacrucian SALMA)
- ?Genus Eutemnodus
- Genus Acrocyon
- Family Proborhyaenidae
- Genus Arminiheringia
- Genus Callistoe
- Genus Paraborhyaena
- Genus Proborhyaena
- Subfamily Thylacosmilinae
- Genus Eomakhaira
- Eomakhaira molossus (early Oligocene, ?Tinguirirican SALMA)[67]
- Genus Anachlysictis
- Genus Patagosmilus[76]
- Patagosmilus goini (early middle Miocene, Colloncuran SALMA)
- Genus Thylacosmilus
- Thylacosmilus atrox (latest Miocene to late Pliocene, Huayquerian-Chapadmalalan SALMA)
- Genus Eomakhaira
- Genus Angelocabrerus
References
- Guillermo W. Rougier, John R. Wible and Michael J. Novacek. First Implications of Deltatheridium specimens for early marsupial history, Nature 396, 459-463(3 December 1998)
- Bi, S.; Jin, X.; Li, S.; Du, T. (2015). "A new Cretaceous metatherian mammal from Henan, China". PeerJ. 3: e896. doi:10.7717/peerj.896. PMC 4400878. PMID 25893149.
- Guillermo Rougier, New specimen of Deltatheroides cretacicus (Metatheria, Deltatheroida) from the Late Cretaceous of Mongolia, BULLETIN OF CARNEGIE MUSEUM OF NATURAL HISTORY 36(DEC 2004):245-266 · SEPTEMBER 2009
- Rougier, Guillermo W.; Davis, Brian M.; Novacek, Michael J. (2015). "A deltatheroidan mammal from the Upper Cretaceous Baynshiree Formation, eastern Mongolia". Cretaceous Research. 52: 167–177. doi:10.1016/j.cretres.2014.09.009.
- Wilson, GP; Ekdale, EG; Hoganson, JW; Calede, JJ; Vander Linden, A (2016). "A large carnivorous mammal from the Late Cretaceous and the North American origin of marsupials". Nat Commun. 7: 13734. doi:10.1038/ncomms13734. PMC 5155139. PMID 27929063.
- Forasiepi, Analía M.; Agustín G. Martinelli; Francisco J. Goín (2007). "Revisión taxonómica de Parahyaenodon argentinus Ameghino y sus implicancias en el conocimiento de los grandes mamíferos carnívoros del Mio-Plioceno de América de Sur". Ameghiniana. 44 (1): 143–159.
- Prevosti, Francisco J.; Analía Forasiepi; Natalia Zimicz (2013). "The Evolution of the Cenozoic Terrestrial Mammalian Predator Guild in South America: Competition or Replacement?". Journal of Mammalian Evolution. 20: 3–21. doi:10.1007/s10914-011-9175-9. S2CID 15751319.
- Engelman, Russell K.; Croft, Darin A. (2019). "Strangers in a strange land: Ecological dissimilarity to metatherian carnivores may partly explain early colonization of South America by Cyonasua-group procyonids". Paleobiology. 45 (4): 598–611. doi:10.1017/pab.2019.29.
- Simpson, George Gaylord (1941). "The affinities of the Borhyaenidae". American Museum Novitates. 1118: 1–6.
- Naish, Darren. "Invasion of the marsupial weasels, dogs, cats and bears...or is it?". Tetrapod Zoology. Scientific American. Retrieved 15 June 2016.
- Argot, Christine (April 2004). "Evolution of South American mammalian predators (Borhyaenoidea): anatomical and palaeobiological implications". Zoological Journal of the Linnean Society. 140 (4): 487–521. doi:10.1111/j.1096-3642.2004.00110.x.
- Forasiepi, Analía M. (2009). "Osteology of Arctodictis sinclairi (Mammalia, Metatheria, Sparassodonta) and phylogeny of Cenozoic metatherian carnivores from South America". Monografías del Museo Argentino de Ciencias Naturales. 6: 1–174.
- Marshall, Larry G.; J.A. Case; M.O. Woodburne (1990). "Phylogenetic Relationships of the Families of Marsupials". Current Mammalogy. 2: 433–505.
- Ercoli, Marcos D.; Francisco J. Prevosti (2011). "Estimación de masa de las especies de Sparassodonta (Mammalia, Metatheria) de edad Santacrucense (Mioceno Temprano) a partir del tamaño del centroide de los elementos apendiculares: inferencias paleoecológicas". Ameghiniana. 48 (4): 462–479. doi:10.5710/amgh.v48i4(347). S2CID 129838311.
- Christian de Muizon; Sandrine Ladevèze; Charlène Selva; Robin Vignaud; Florent Goussard (2018). "Allqokirus australis (Sparassodonta, Metatheria) from the early Palaeocene of Tiupampa (Bolivia) and the rise of the metatherian carnivorous radiation in South America". Geodiversitas. 40 (16): 363–459. doi:10.5252/geodiversitas2018v40a16.
- Shockey, Bruce J.; Anaya, Federico (2008). "Postcranial Osteology of Mammals from Salla, Bolivia (Late Oligocene): Form, Function, and Phylogenetic Implications". In Eric J. Sargis; Dagosto, Marian (eds.). Mammalian Evolutionary Morphology: a Tribute to Frederick S. Szalay. Springer. pp. 135–157. ISBN 978-1-4020-6997-0.
- Marshall, Larry G. (1978). Evolution of the Borhyaenidae, extinct South American predaceous marsupials. University of California Publications in Geological Sciences. 117. pp. 1–89. ISBN 9780520095717.
- Marshall, Larry G. (1976). "Notes on the deciduous dentition of the Borhyaenidae (Marsupialia: Borhyaenoidea)". Journal of Mammalogy. 57 (4): 751–754. doi:10.2307/1379446. JSTOR 1379446.
- Forasiepi, Analia M.; Marcelo R. Sanchez-Villagra (2014). "Heterochrony, dental ontogenetic diversity, and the circumvention of constraints in marsupial mammals and extinct relatives". Paleobiology. 40 (2): 222–237. doi:10.1666/13034. S2CID 84605309.
- Wm. J. Sinclair, The Marsupial Fauna of the Santa Cruz Beds, Proceedings of the American Philosophical Society . Vol. 44, No. 179 (Jan. - Apr., 1905), pp. 73-81 . Published by: American Philosophical Society
- Florentino, Ameghino (1887). "Enumeración sistemática de las especies de mamíferos fósiles coleccionados for Carlos Ameghino en los terrenos Eocenos de Patagonia Austral". Boletim Museo la Plata. 1: 1–26.
- Matthew, W. D. (1907). "II.—The Relationships of the 'Sparassodonta.'". Geological Magazine. 4 (12): 531–535. doi:10.1017/S0016756800134090.
- Sinclair, William J. (1906). "Mammalia of the Santa Cruz Beds. Marsupialia". Reports of the Princeton University Expedition to Patagonia. 4: 333–460.
- Wood, Horace Elmer (1924). "The position of the "sparassodonts": with notes on the relationships and history of the Marsupialia". Bulletin of the American Museum of Natural History. 51 (4): 77–101.
- Cabrera, Ángel (1927). "Datos para el conocimiento de los dasiuroideos fósiles argentinos". Revista del Museo de la Plata. 30: 271–315.
- Marshall, L. G. (1 December 1977). "Cladistic Analysis of Borhyaenoid, Dasyuroid, Didelphoid, and Thylacinid (Marsupialia: Mammalia) Affinity". Systematic Biology. 26 (4): 410–425. doi:10.1093/sysbio/26.4.410.
- Marshall, Larry G.; Kielan-Jaworowska, Zofia (October 1992). "Relationships of the dog-like marsupials, deltatheroidans and early tribosphenic mammals". Lethaia. 25 (4): 361–374. doi:10.1111/j.1502-3931.1992.tb01639.x.
- Muizon, Christian; Lange-Badré, Brigitte (1997). "Carnivorous dental adaptations in tribosphenic mammals and phylogenetic reconstruction". Lethaia. 30 (4): 353–366. doi:10.1111/j.1502-3931.1997.tb00481.x.
- Szalay, Frederick S. (1994). Evolutionary history of the marsupials and an analysis of osteological characters. Cambridge: Cambridge University Press. pp. 1–496. ISBN 978-0521025928.
- Rangel, Caio C.; Carneiro, Leonardo M.; Bergqvist, Lilian P.; Oliveira, Edison V.; Goin, Francisco J.; Babot, Maria J. (2019). "Diversity, affinities, and Adaptations of the Basal Sparassodont Patene Simpson, 1935 (Mammalia, Metatheria)". Ameghiniana. 56: 263. doi:10.5710/AMGH.06.05.2019.3222.
- Beck, R. M. D. (2013). "A peculiar faunivorous metatherian from the early Eocene of Australia". Acta Palaeontologica Polonica. 60 (1): 123–129.
- Leonardo M. Carneiro (2018). "A new species of Varalphadon (Mammalia, Metatheria, Sparassodonta) from the upper Cenomanian of southern Utah, North America: Phylogenetic and biogeographic insights". Cretaceous Research. 84: 88–96. doi:10.1016/j.cretres.2017.11.004.
- Goin, Francisco J.; Vieytes, Emma C.; Gelfo, Javier N.; Chornogubsky, Laura; Zimicz, Ana N.; Reguero, Marcelo A. (20 September 2018). "New Metatherian Mammal from the Early Eocene of Antarctica". Journal of Mammalian Evolution. 27: 17–36. doi:10.1007/s10914-018-9449-6. S2CID 91932037.
- Babot, María J.; Jaime E. Powell; Christian de Muizon (2002). "Callistoe vincei a new Proborhyaenidae (Borhyaenoidea, Metatheria, Mammalia) from the Early Eocene of Argentina". Geobios. 35 (5): 615–629. doi:10.1016/S0016-6995(02)00073-6.
- Wilson, G.P.; Ekdale, E.G.; Hoganson, J.W.; Calede, J.J.; Linden, A.V. (2016). "A large carnivorous mammal from the Late Cretaceous and the North American origin of marsupials". Nature Communications. 7: 13734. doi:10.1038/ncomms13734. PMC 5155139. PMID 27929063.
- Forasiepi, Analía M.; Guillermo. W. Rougier (2009). "Additional data on early Paleocene metatherians (Mammalia) from Punta Peligro (Salamanca Formation, Argentina): comments based on petrosal morphology". Journal of Zoological Systematics and Evolutionary Research. 47 (4): 391–398. doi:10.1111/j.1439-0469.2008.00519.x.
- Babot, M. Judith; Pablo E. Ortiz (2008). "Primer registro de Borhyaenoidea (Mammalia, Metatheria, Sparassodonta) en la provincia de Tucumán (Formacion India Muerta, Grupo Choromoro; Mioceno tardío)". Acta Geológica Lilloana. 21 (1): 34–48.
- Goin, Francisco J.; Ulyses F. J. Pardiñas (1996). "Revision de las especies del genero Hyperdidelphys Ameghino, 1904 (Mammalia, Marsupialia, Didelphidae. Su significacion filogenetica, estratigrafica y adaptativa en el Neogeno del Cono Sur sudamericano". Estudios Geologicos. 52 (5–6): 327–359. doi:10.3989/egeol.96525-6275.
- Prevosti, Francisco J.; Ulyses F. J. Pardinas (2009). "Comment on "The oldest South American Cricetidae (Rodentia) and Mustelidae (Carnivora): Late Miocene faunal turnover in central Argentina and the Great American Biotic Interchange" by D.H. Verzi and C.I. Montalvo [Palaeogeography, Palaeoclimatology, Palaeoecology 267 (2008) 284–291]". Palaeogeography, Palaeoclimatology, Palaeoecology. 280 (3–4): 543–547. doi:10.1016/j.palaeo.2009.05.021.
- Zimicz, Natalia (2014). "Avoiding Competition: the Ecological History of Late Cenozoic Metatherian Carnivores in South America". Journal of Mammalian Evolution. 21 (4): 383–393. doi:10.1007/s10914-014-9255-8. S2CID 10161199.
- Croft, Darin A.; Engelman, Russell K.; Dolgushina, Tatiana; Wesley, Gina (2018). "Diversity and disparity of sparassodonts (Metatheria) reveal non-analogue nature of ancient South American mammalian carnivore guilds". Proceedings of the Royal Society B: Biological Sciences. 285 (1870): 20172012. doi:10.1098/rspb.2017.2012. PMC 5784193. PMID 29298933.
- Marshall, Larry G. (1977). "A New Species of Lycopsis (Borhyaenidae: Marsupialia) from the La Venta Fauna (Late Miocene) of Colombia, South America". Journal of Paleontology. 51 (3): 633–642. ISSN 0022-3360. JSTOR 1303691.
- Tomassini, Rodrigo L.; Montalvo, Claudia I.; Bargo, M. Susana; Vizcaíno, Sergio F.; Cuitiño, José I. (23 December 2019). "Sparassodonta (Metatheria) coprolites from the early-mid Miocene (Santacrucian age) of Patagonia (Argentina) with evidence of exploitation by coprophagous insects". PALAIOS. 34 (12): 639–651. doi:10.2110/palo.2019.080. S2CID 209473467.
- Tomassini, Rodrigo L.; Garrone, Mariana C.; Montalvo, Claudia I. (2017). "New light on the endemic South American pachyrukhine Paedotherium Burmeister, 1888 (Notoungulata, Hegetotheriidae): Taphonomic and paleohistological analysis". Journal of South American Earth Sciences. 73: 33–41. doi:10.1016/j.jsames.2016.11.004.
- Domingo, Laura; Tomassini, Rodrigo L.; Montalvo, Claudia I.; Sanz-Pérez, Dánae; Alberdi, María Teresa (December 2020). "The Great American Biotic Interchange revisited: a new perspective from the stable isotope record of Argentine Pampas fossil mammals". Scientific Reports. 10 (1): 1608. doi:10.1038/s41598-020-58575-6. ISSN 2045-2322. PMC 6994648. PMID 32005879.
- Cione, Alberto Luis; Acosta Hospitaleche, Carolina; Pérez, Leandro Martín; Laza, Jose Herminio; César, Inés (2010). "Trace fossils on penguin bones from the Miocene of Chubut, southern Argentina". Alcheringa: An Australasian Journal of Palaeontology. 34 (4): 433–454. doi:10.1080/03115511003793470. S2CID 140681677.
- Mendoza, Ricardo S. De; Haidr, Nadia S. (October 2018). "Predation Trace Fossils in a New Specimen of Cayaoa bruneti Tonni (Aves, Anseriformes) from the Gaiman Formation (Early Miocene, Chubut, Argentina)". Ameghiniana. 55 (4): 483–488. doi:10.5710/AMGH.02.02.2018.3111. S2CID 134488773.
- Blanco, R. Ernesto; Jones, Washington W.; Grinspan, Gustavo A. (September 2011). "Fossil marsupial predators of South America (Marsupialia, Borhyaenoidea): bite mechanics and palaeobiological implications". Alcheringa: An Australasian Journal of Palaeontology. 35 (3): 377–387. doi:10.1080/03115518.2010.519644. S2CID 85017417.
- Ercoli, Marcos D.; Prevosti, Francisco J.; Forasiepi, Analía M. (8 October 2013). "The Structure of the Mammalian Predator Guild in the Santa Cruz Formation (Late Early Miocene)" (PDF). Journal of Mammalian Evolution. 21 (4): 369–381. doi:10.1007/s10914-013-9243-4. S2CID 6240294.
- Echarri, Sebastian; Ercoli, Marcos D.; Amelia Chemisquy, M.; Turazzini, Guillermo; Prevosti, Francisco J. (16 January 2017). "Mandible morphology and diet of the South American extinct metatherian predators (Mammalia, Metatheria, Sparassodonta)". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 106 (4): 277–288. doi:10.1017/S1755691016000190.
- Analía M. Forasiepi, M. Judith Babot and Natalia Zimicz (2014). "Australohyaena antiqua (Mammalia, Metatheria, Sparassodonta), a large predator from the Late Oligocene of Patagonia". Journal of Systematic Palaeontology. 13 (6): 1–23. doi:10.1080/14772019.2014.926403. S2CID 83669335.
- Argot, Christine (2004). "Functional-adaptive analysis of the postcranial skeleton of a Laventan borhyaenoid, Lycopsis longirostris (Marsupialia, Metatheria)". Journal of Vertebrate Paleontology. 24 (3): 689–708. doi:10.1671/0272-4634(2004)024[0689:FAOTPS]2.0.CO;2.
- Ercoli, Marcos D.; Francisco J. Prevosti; Alicia Álvarez (2012). "Form and function within a phylogenetic framework: locomotory habits of extant predators and some Miocene Sparassodonta". Zoological Journal of the Linnean Society. 165: 224–251. doi:10.1111/j.1096-3642.2011.00793.x.
- Argot, Christine; Judith Babot (2011). "Postcranial morphology, functional adaptations and paleobiology of Callistoe vincei, a predaceous metatherian from the Eocene of Salta, north-western Argentina". Palaeontology. 54 (2): 447–480. doi:10.1111/j.1475-4983.2011.01036.x.
- Riggs, Elmer S. (1934). "A New Marsupial Saber-Tooth from the Pliocene of Argentina and Its Relationships to Other South American Predacious Marsupials". Transactions of the American Philosophical Society. 24 (1): 1–32. doi:10.2307/3231954. JSTOR 3231954.
- Argot, Christine (January 2004). "Functional-adaptive features and palaeobiologic implications of the postcranial skeleton of the late Miocene sabretooth borhyaenoid Thylacosmilus atrox (Metatheria)". Alcheringa: An Australasian Journal of Palaeontology. 28 (1): 229–266. doi:10.1080/03115510408619283. S2CID 129220124.
- Aramayo, Silvia A. (2007). "Neogene vertebrate palaeoichnology of the North Atlantic coast of the Rio Negro Province, Argentina". Arquivos do Museu Nacional Rio de Janeiro. 65 (4): 573–584.
- Cladera, Gerardo; Ruigomez, Eduardo; Jaureguizar, Edgardo Ortíz; Bond, Mariano; López, Guillermo (2004). "Tafonomía de la Gran Hondonada (Formación Sarmiento, Edad-mamífero Mustersense, Eoceno Medio) Chubut, Argentina". Ameghiniana (in Spanish). 41 (3): 315–330. ISSN 1851-8044.
- Croft, Darin A. (2006). "Do marsupials make good predators? Insights from predator–prey diversity ratios". Evolutionary Ecology Research. 8 (7): 1193–1214. ISSN 1522-0613.
- Vizcaíno, Sergio F.; Bargo, M. Susana; Kay, Richard F.; Fariña, Richard A.; Di Giacomo, Mariana; Perry, Jonathan M.G.; Prevosti, Francisco J.; Toledo, Néstor; Cassini, Guillermo H.; Fernicola, Juan C. (June 2010). "A baseline paleoecological study for the Santa Cruz Formation (late–early Miocene) at the Atlantic coast of Patagonia, Argentina". Palaeogeography, Palaeoclimatology, Palaeoecology. 292 (3–4): 507–519. doi:10.1016/j.palaeo.2010.04.022.
- Engelman, Russell K.; Anaya, Federico; Croft, Darin A. (2015). "New Specimens of Acyon myctoderos (Metatheria, Sparassodonta) from Quebrada Honda, Bolivia". Ameghiniana. 52 (2): 204. doi:10.5710/AMGH.19.11.2014.2803. S2CID 128596671.
- Dozo, María Teresa (1994). "Estudios paleoneurologicos en marsupiales "carnivoros" extinguidos de America del Sur: Neuromorphologia y encefalizacion". Mastozoología Neotropical. 1 (1): 5–16.
- Ercoli, Marcos Darío; Prevosti, Francisco Juan (1 December 2011). "Estimación de Masa de las Especies de Sparassodonta (Mammalia, Metatheria) de Edad Santacrucense (Mioceno Temprano) a Partir del Tamaño del Centroide de los Elementos Apendiculares: Inferencias Paleoecológicas". Ameghiniana. 48 (4): 462. doi:10.5710/AMGH.v48i4(347). S2CID 129838311.
- Savage, R.J.G. (1977). "Evolution in carnivorous mammals". Palaeontology. 20 (2): 237–271.
- Forasiepi, Analía M.; Macphee, Ross D.E.; Pino, Santiago Hernández del (14 June 2019). "Caudal Cranium of Thylacosmilus atrox (Mammalia, Metatheria, Sparassodonta), a South American Predaceous Sabertooth". Bulletin of the American Museum of Natural History. 2019 (433): 1. doi:10.1206/0003-0090.433.1.1.
- Goin, Francisco J.; Abello, María Alejandra (1 February 2013). "Los Metatheria Sudamericanos de Comienzos Del Neógeno (Mioceno Temprano, Edad MamÍFero Colhuehuapense): Microbiotheria y Polydolopimorphia". Ameghiniana. 50 (1): 51. doi:10.5710/AMGH.9.11.2012.570. hdl:11336/76812. S2CID 129111824.
- Engelman, Russell K.; Flynn, John J.; Wyss, André R.; Croft, Darin A. (17 July 2020). "Eomakhaira molossus, A New Saber-Toothed Sparassodont (Metatheria: Thylacosmilinae) from the Early Oligocene (?Tinguirirican) Cachapoal Locality, Andean Main Range, Chile". American Museum Novitates. 2020 (3957): 1. doi:10.1206/3957.1. S2CID 220601822.
- Suárez, C.; Forasiepi, A. M.; Goin, F. J.; Jaramillo, C. (2015). "Insights into the Neotropics prior to the Great American Biotic Interchange: new evidence of mammalian predators from the Miocene of Northern Colombia". Journal of Vertebrate Paleontology. Online edition: e1029581. doi:10.1080/02724634.2015.1029581. S2CID 86264178.
- Wilson, G.P.; Ekdale, E.G.; Hoganson, J.W.; Calede, J.J.; Linden, A.V. (2016). "A large carnivorous mammal from the Late Cretaceous and the North American origin of marsupials". Nature Communications. 7: 13734. doi:10.1038/ncomms13734. PMC 5155139. PMID 27929063.
- Prevosti, Francisco J. Evolution of South American Mammalian Predators During the Cenozoic: Paleobiogeographic and Paleoenvironmental Contingencies. Cham: Springer. pp. 1–196.
- de Muizon, Christian (1998). "Mayulestes ferox, a borhyaenoid (Metatheria, Mammalia) from the early Paleocene of Bolivia. Phylogenetic and palaeobiologic implications". Geodiversitas. 20: 19–142.
- Forasiepi, Analía M.; Marcelo R. Sánchez-Villagra, Marcelo R.; Francisco J. Goin, Francisco J.; Masanaru Takai, Masanaru; Nobuo Shigehara, Nobuo; Richard F. Kay, Richard F. (2006). "A new species of Hathliacynidae (Metatheria, Sparassodonta) from the middle Miocene of Quebrada Honda, Bolivia". Journal of Vertebrate Paleontology. 26 (3): 670–684. doi:10.1671/0272-4634(2006)26[670:ANSOHM]2.0.CO;2.
- de Muizon, Christian (1999). "Marsupial skulls from the Deseadan (late Oligocene) of Bolivia and phylogenetic analysis of the Borhyaenoidea". Geobios. 32 (3): 483–509. doi:10.1016/s0016-6995(99)80022-9.
- Engelman, Russell K.; Anaya, Federico; Croft, Darin A. (2018). "Australogale leptognathus, gen. et sp. nov., a Second Species of Small Sparassodont (Mammalia: Metatheria) from the Middle Miocene Locality of Quebrada Honda, Bolivia". Journal of Mammalian Evolution. 27: 1–18. doi:10.1007/s10914-018-9443-z. S2CID 49473591.
- Engelman, Russell K.; Flynn, John J.; Gans, Philip; Wyss, André R.; Croft, Darin (2018). "Chlorocyon phantasma, a Late Eocene Borhyaenoid (Mammalia: Metatheria: Sparassodonta) from the Los Helados Locality, Andean Main Range, Central Chile". American Museum Novitates. 3918 (3918): 1–23. doi:10.1206/3918.1. S2CID 92580823.
- Forasiepi, Analía M.; Alfredo A. Carlini (2010). "A new thylacosmilid (Mammalia, Metatheria, Sparassodonta) from the Miocene of Patagonia, Argentina" (PDF). Zootaxa. 2552: 55–68. doi:10.11646/zootaxa.2552.1.3.
Further reading
- Goin, Francisco J (2003). "Early Marsupial radiations in South America". In Menna Jones; Mike Archer; Chris Dickman (eds.). Predators with Pouches: The Biology of Carnivorous Marsupials. CSIRO. pp. 30–42. ISBN 9780643099487.
External links
| Wikimedia Commons has media related to Sparassodonta. |
| Wikispecies has information related to Sparassodonta. |
| Wikisource has the text of the 1911 Encyclopædia Britannica article Sparassodonta. |
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||




