Biological pump
The biological pump, also known as the marine carbon pump, is, in its simplest form, the ocean's biologically driven sequestration of carbon from the atmosphere and land runoff to the ocean interior and seafloor sediments.[1] It is the part of the oceanic carbon cycle responsible for the cycling of organic matter formed mainly by phytoplankton during photosynthesis (soft-tissue pump), as well as the cycling of calcium carbonate (CaCO3) formed into shells by certain organisms such as plankton and mollusks (carbonate pump).[2]
| Part of a series of overviews on |
| Marine life |
|---|
|
|
The biological pump is not so much the result of a single process, but rather the sum of a number of processes each of which can influence biological pumping.
Overview
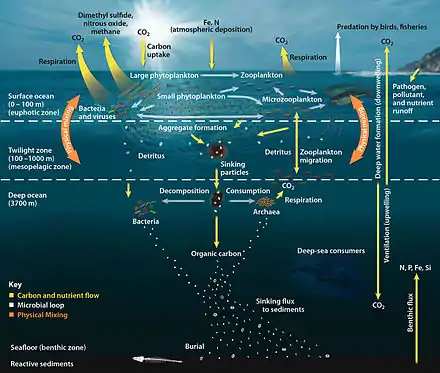
The biological pump can be divided into three distinct phases,[4] the first of which is the production of fixed carbon by planktonic phototrophs in the euphotic (sunlit) surface region of the ocean. In these surface waters, phytoplankton use carbon dioxide (CO2), nitrogen (N), phosphorus (P), and other trace elements (barium, iron, zinc, etc.) during photosynthesis to make carbohydrates, lipids, and proteins. Some plankton, (e.g. coccolithophores and foraminifera) combine calcium (Ca) and dissolved carbonates (carbonic acid and bicarbonate) to form a calcium carbonate (CaCO3) protective coating.
Once this carbon is fixed into soft or hard tissue, the organisms either stay in the euphotic zone to be recycled as part of the regenerative nutrient cycle or once they die, continue to the second phase of the biological pump and begin to sink to the ocean floor. The sinking particles will often form aggregates as they sink, greatly increasing the sinking rate. It is this aggregation that gives particles a better chance of escaping predation and decomposition in the water column and eventually making it to the sea floor.
The fixed carbon that is decomposed by bacteria either on the way down or once on the sea floor then enters the final phase of the pump and is remineralized to be used again in primary production. The particles that escape these processes entirely are sequestered in the sediment and may remain there for millions of years. It is this sequestered carbon that is responsible for ultimately lowering atmospheric CO2.
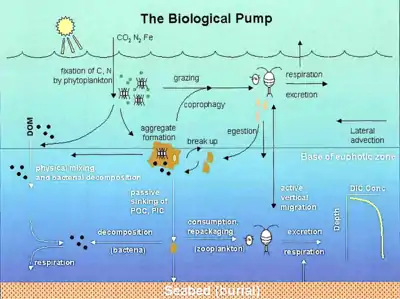
The biological pump is responsible for transforming dissolved inorganic carbon (DIC) into organic biomass and pumping it in particulate or dissolved form into the deep ocean. Inorganic nutrients and carbon dioxide are fixed during photosynthesis by phytoplankton, which both release dissolved organic matter (DOM) and are consumed by herbivorous zooplankton. Larger zooplankton - such as copepods - egest fecal pellets which can be reingested and sink or collect with other organic detritus into larger, more-rapidly-sinking aggregates. DOM is partially consumed by bacteria (black dots) and respired; the remaining refractory DOM is advected and mixed into the deep sea. DOM and aggregates exported into the deep water are consumed and respired, thus returning organic carbon into the enormous deep ocean reservoir of DIC. About 1% of the particles leaving the surface ocean reach the seabed and are consumed, respired, or buried in the sediments. There, carbon is stored for millions of years. The net effect of these processes is to remove carbon in organic form from the surface and return it to DIC at greater depths, maintaining the surface-to-deep ocean gradient of DIC. Thermohaline circulation returns deep-ocean DIC to the atmosphere on millennial timescales.[5]
Primary production
| Part of a series on the |
| Carbon cycle |
|---|
 |
The first step in the biological pump is the synthesis of both organic and inorganic carbon compounds by phytoplankton in the uppermost, sunlit layers of the ocean.[8] Organic compounds in the form of sugars, carbohydrates, lipids, and proteins are synthesized during the process of photosynthesis:
CO2 + H2O + light → CH2O + O2
In addition to carbon, organic matter found in phytoplankton is composed of nitrogen, phosphorus and various trace metals. The ratio of carbon to nitrogen and phosphorus varies from place to place,[9] but has an average ratio near 106C:16N:1P, known as the Redfield ratio. Trace metals such as magnesium, cadmium, iron, calcium, barium and copper are orders of magnitude less prevalent in phytoplankton organic material, but necessary for certain metabolic processes and therefore can be limiting nutrients in photosynthesis due to their lower abundance in the water column.[4]
Oceanic primary production accounts for about half of the carbon fixation carried out on Earth. Approximately 50–60 Pg of carbon are fixed by marine phytoplankton each year despite the fact that they comprise less than 1% of the total photosynthetic biomass on Earth. The majority of this carbon fixation (~80%) is carried out in the open ocean while the remaining amount occurs in the very productive upwelling regions of the ocean. Despite these productive regions producing 2 to 3 times as much fixed carbon per area, the open ocean accounts for greater than 90% of the ocean area and therefore is the larger contributor.[4]
Calcium carbonate

Carbon is also biologically fixed in the form of calcium carbonate (CaCO3) used as a protective coating for many planktonic species (coccolithophores, foraminifera) as well as larger marine organisms (mollusk shells). Calcium carbonate is also excreted at high rates by fish.[10] While this form of carbon is not directly taken from the atmospheric budget, it is formed from dissolved forms of carbonate which are in equilibrium with CO2 and then responsible for removing this carbon via sequestration.[11]
CO2 + H2O → H2CO3 → H+ + HCO3−
Ca2+ + 2HCO3− → CaCO3 + CO2 + H2O
While this process does manage to fix a large amount of carbon, two units of alkalinity are sequestered for every unit of sequestered carbon.[2][12] The formation and sinking of CaCO3 therefore drives a surface to deep alkalinity gradient which serves to raise the pH of surface waters, shifting the speciation of dissolved carbon to raise the partial pressure of dissolved CO2 in surface waters, which actually raises atmospheric levels. In addition, the burial of CaCO3 in sediments serves to lower overall oceanic alkalinity, tending to raise pH and thereby atmospheric CO2 levels if not counterbalanced by the new input of alkalinity from weathering.[1] The portion of carbon that is permanently buried at the sea floor becomes part of the geologic record. Calcium carbonate often forms remarkable deposits that can then be raised onto land through tectonic motion as in the case with the White Cliffs of Dover in Southern England. These cliffs are made almost entirely of the plates of buried coccolithophores.
Oceanic carbon cycle
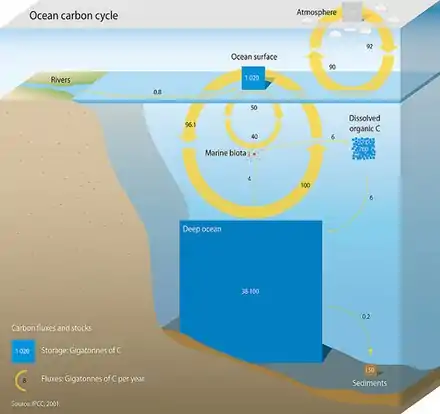
Three main processes (or pumps) that make up the marine carbon cycle bring atmospheric carbon dioxide (CO2) into the ocean interior and distribute it through the oceans. These three pumps are: (1) the solubility pump, (2) the carbonate pump, and (3) the biological pump. The total active pool of carbon at the Earth's surface for durations of less than 10,000 years is roughly 40,000 gigatons C (Gt C, a gigaton is one billion tons, or the weight of approximately 6 million blue whales), and about 95% (~38,000 Gt C) is stored in the ocean, mostly as dissolved inorganic carbon.[13][14] The speciation of dissolved inorganic carbon in the marine carbon cycle is a primary controller of acid-base chemistry in the oceans.
Solubility pump
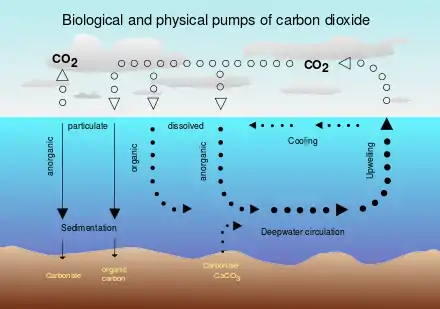
The biological pump is accompanied by a physico-chemical counterpart known as the solubility pump. This pump transports significant amounts of carbon in the form of dissolved inorganic carbon (DIC) from the ocean's surface to its interior. It involves physical and chemical processes only, and does not involve biological processes.[15]
The solubility pump is driven by the coincidence of two processes in the ocean:
- The solubility of carbon dioxide is a strong inverse function of seawater temperature (i.e. solubility is greater in cooler water)
- The thermohaline circulation is driven by the formation of deep water at high latitudes where seawater is usually cooler and denser
Since deep water (that is, seawater in the ocean's interior) is formed under the same surface conditions that promote carbon dioxide solubility, it contains a higher concentration of dissolved inorganic carbon than might be expected from average surface concentrations. Consequently, these two processes act together to pump carbon from the atmosphere into the ocean's interior. One consequence of this is that when deep water upwells in warmer, equatorial latitudes, it strongly outgasses carbon dioxide to the atmosphere because of the reduced solubility of the gas.[16]
Carbonate pump

The carbonate pump is sometimes referred to as the “hard tissue” component of the biological pump.[17] Some surface marine organisms, like Coccolithophores, produce hard structures out of calcium carbonate, a form of particulate inorganic carbon, by fixing bicarbonate.[18] This fixation of DIC is an important part of the oceanic carbon cycle.
Ca2+ + 2 HCO3− → CaCO3 + CO2 + H2O
While the biological carbon pump fixes inorganic carbon (CO2) into particulate organic carbon in the form of sugar (C6H12O6), the carbonate pump fixes inorganic bicarbonate and causes a net release of CO2.[18] In this way, the carbonate pump could be termed the carbonate counter pump. It works counter to the biological pump by counteracting the CO2 flux from the biological pump.
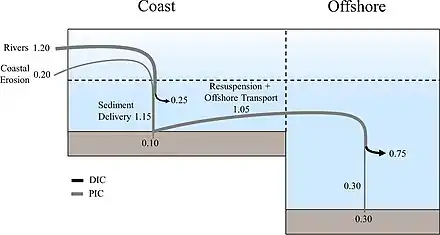
Continental shelf pump
The continental shelf pump is proposed as operating in the shallow waters of the continental shelves as a mechanism transporting carbon (dissolved or particulate) from the continental waters to the interior of the adjacent deep ocean.[19] As originally formulated, the pump is thought to occur where the solubility pump interacts with cooler, and therefore denser water from the shelf floor which feeds down the continental slope into the neighbouring deep ocean.[19] The shallowness of the continental shelf restricts the convection of cooling water, so the cooling can be greater for continental shelf waters than for neighbouring open ocean waters. These cooler waters promote the solubility pump and lead to an increased storage of dissolved inorganic carbon. This extra carbon storage is further augmented by the increased biological production characteristic of shelves.[20] The dense, carbon-rich shelf waters then sink to the shelf floor and enter the sub-surface layer of the open ocean via isopycnal mixing.[19] As the sea level rises in response to global warming, the surface area of the shelf seas will grow and in consequence the strength of the shelf sea pump should increase.[21]
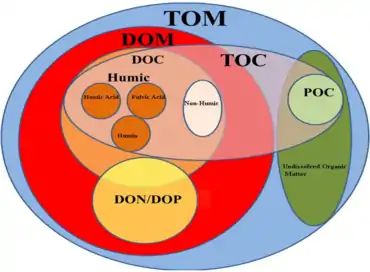
Types of carbon
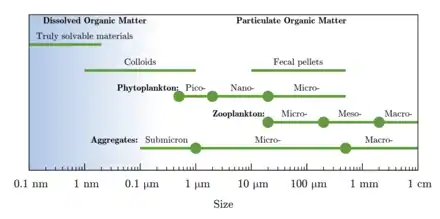
- Dissolved and particulate carbon
Phytoplankton supports all life in the ocean as it converts inorganic compounds into organic constituents. This autotrophically produced biomass presents the foundation of the marine food web.[22]
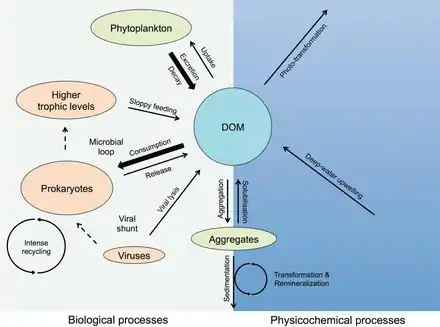
Processes in the biological pump
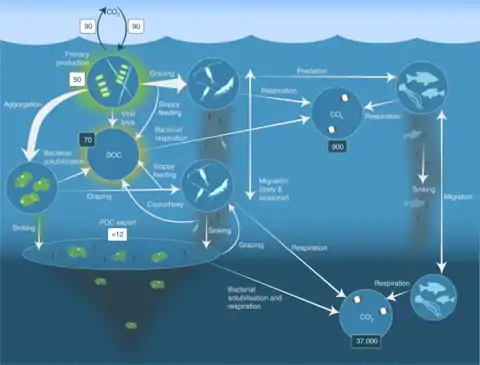
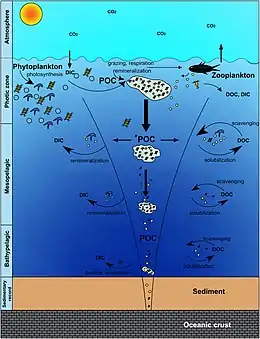
In the diagram on the right, phytoplankton convert CO2, which has dissolved from the atmosphere into the surface oceans (90 Gt yr−1), into particulate organic carbon (POC) during primary production (~ 50 Gt C yr−1). Phytoplankton are then consumed by copepods, krill and other small zooplankton grazers, which in turn are preyed upon by higher trophic levels. Any unconsumed phytoplankton form aggregates, and along with zooplankton faecal pellets, sink rapidly and are exported out of the mixed layer (< 12 Gt C yr−1 14). Krill, copepods, zooplankton and microbes intercept phytoplankton in the surface ocean and sinking detrital particles at depth, consuming and respiring this POC to CO2 (dissolved inorganic carbon, DIC), such that only a small proportion of surface-produced carbon sinks to the deep ocean (i.e., depths > 1000 m). As krill and smaller zooplankton feed, they also physically fragment particles into small, slower- or non-sinking pieces (via sloppy feeding, coprorhexy if fragmenting faeces),[26] retarding POC export. This releases dissolved organic carbon (DOC) either directly from cells or indirectly via bacterial solubilisation (yellow circle around DOC). Bacteria can then remineralise the DOC to DIC (CO2, microbial gardening).[25]
Marine snow
Most carbon incorporated in organic and inorganic biological matter is formed at the sea surface where it can then start sinking to the ocean floor. The deep ocean gets most of its nutrients from the higher water column when they sink down in the form of marine snow. This is made up of dead or dying animals and microbes, fecal matter, sand and other inorganic material.[27] A single phytoplankton cell has a sinking rate around one metre per day. Given that the average depth of the ocean is about four kilometres, it can take over ten years for these cells to reach the ocean floor. However, through processes such as coagulation and expulsion in predator fecal pellets, these cells form aggregates. These aggregates, known as marine snow, have sinking rates orders of magnitude greater than individual cells and complete their journey to the deep in a matter of days.[4]
Of the 50–60 Pg of carbon fixed annually, roughly 10% leaves the surface mixed layer of the oceans, while less than 0.5% of eventually reaches the sea floor.[4] Most is retained in regenerated production in the euphotic zone and a significant portion is remineralized in midwater processes during particle sinking. The portion of carbon that leaves the surface mixed layer of the ocean is sometimes considered "sequestered", and essentially removed from contact with the atmosphere for many centuries.[28] However, work also finds that, in regions such as the Southern Ocean, much of this carbon can quickly (within decades) come back into contact with the atmosphere.[29]
Sloppy feeding
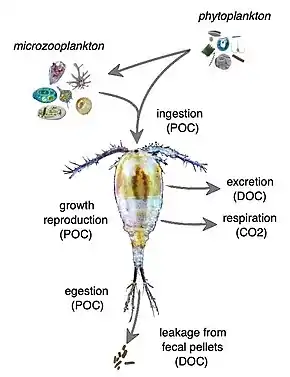
In addition to linking primary producers to higher trophic levels in marine food webs, zooplankton also play an important role as “recyclers” of carbon and other nutrients that significantly impact marine biogeochemical cycles, including the biological pump. This is particularly the case with copepods and krill, and is especially important in oligotrophic waters of the open ocean. Through sloppy feeding, excretion, egestion, and leaching of fecal pellets, zooplankton release dissolved organic matter (DOM) which controls DOM cycling and supports the microbial loop. Absorption efficiency, respiration, and prey size all further complicate how zooplankton are able to transform and deliver carbon to the deep ocean.[32]
Excretion and sloppy feeding (the physical breakdown of food source) make up 80% and 20% of crustacean zooplankton-mediated DOM release respectively.[33] In the same study, fecal pellet leaching was found to be an insignificant contributor. For protozoan grazers, DOM is released primarily through excretion and egestion and gelatinous zooplankton can also release DOM through the production of mucus. Leaching of fecal pellets can extend from hours to days after initial egestion and its effects can vary depending on food concentration and quality.[34][35] Various factors can affect how much DOM is released from zooplankton individuals or populations.
Fecal pellets
Absorption efficiency (AE) is the proportion of food absorbed by plankton that determines how available the consumed organic materials are in meeting the required physiological demands.[32] Depending on the feeding rate and prey composition, variations in AE may lead to variations in fecal pellet production, and thus regulates how much organic material is recycled back to the marine environment. Low feeding rates typically lead to high AE and small, dense pellets, while high feeding rates typically lead to low AE and larger pellets with more organic content. Another contributing factor to DOM release is respiration rate. Physical factors such as oxygen availability, pH, and light conditions may affect overall oxygen consumption and how much carbon is loss from zooplankton in the form of respired CO2. The relative sizes of zooplankton and prey also mediate how much carbon is released via sloppy feeding. Smaller prey are ingested whole, whereas larger prey may be fed on more “sloppily”, that is more biomatter is released through inefficient consumption.[36][37] There is also evidence that diet composition can impact nutrient release, with carnivorous diets releasing more dissolved organic carbon (DOC) and ammonium than omnivorous diets.[34]
Vertical migrations
Diel vertically migrating krill, salps, smaller zooplankton and fish can actively transport carbon to depth by consuming POC in the surface layer at night, and metabolising it at their daytime, mesopelagic residence depths. Depending on species life history, active transport may occur on a seasonal basis as well.[25]
Without vertical migration the biological pump wouldn't be nearly as efficient. Organisms migrate up to feed at night so when they migrate back to depth during the day they defecate large sinking fecal pellets. Whilst some larger fecal pellets can sink quite fast, the speed that organisms move back to depth is still faster. At night organisms are in the top 100 metres of the water column, but during the day they move down to between 800–1000 meters. If organisms were to defecate at the surface it would take the fecal pellets days to reach the depth that they reach in a matter of hours. Therefore, by releasing fecal pellets at depth they have almost 1000 metres less to travel to get to the deep ocean. This is something known as active transport. The organisms are playing a more active role in moving organic matter down to depths. Because a large majority of the deep sea, especially marine microbes, depends on nutrients falling down, the quicker they can reach the ocean floor the better.[27]
Zooplankton and salps play a large role in the active transport of fecal pellets. 15–50% of zooplankton biomass is estimated to migrate, accounting for the transport of 5–45% of particulate organic nitrogen to depth.[27] Salps are large gelatinous plankton that can vertically migrate 800 meters and eat large amounts of food at the surface. They have a very long gut retention time, so fecal pellets usually are released at maximum depth. Salps are also known for having some of the largest fecal pellets. Because of this they have a very fast sinking rate, small detritus particles are known to aggregate on them. This makes them sink that much faster. So while currently there is still much research being done on why organisms vertically migrate, it is clear that vertical migration plays a large role in the active transport of dissolved organic matter to depth.[38]
Jelly pump

Jelly-falls are marine carbon cycling events whereby gelatinous zooplankton, primarily cnidarians, sink to the seafloor and enhance carbon and nitrogen fluxes via rapidly sinking particulate organic matter.[40] These events provide nutrition to benthic megafauna and bacteria.[41][42] Jelly-falls have been implicated as a major “gelatinous pathway” for the sequestration of labile biogenic carbon through the biological pump.[43] These events are common in protected areas with high levels of primary production and water quality suitable to support cnidarian species. These areas include estuaries and several studies have been conducted in fjords of Norway.[42]
Whale pump
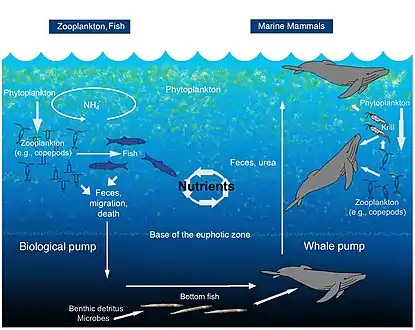
Whales and other marine mammals also enhance primary productivity in their feeding areas by concentrating nitrogen near the surface through the release of flocculent fecal plumes.[44][45] For example, whales and seals may be responsible for replenishing more nitrogen in the Gulf of Maine's euphotic zone than the input of all rivers combined. This upward whale pump played a much larger role before industrial fishing devastated marine mammal stocks, when recycling of nitrogen was likely more than three times the atmospheric nitrogen input.[44]
The biological pump mediates the removal of carbon and nitrogen from the euphotic zone through the downward flux of aggregates, feces, and vertical migration of invertebrates and fish.[46] Copepods and other zooplankton produce sinking fecal pellets and contribute to downward transport of dissolved and particulate organic matter by respiring and excreting at depth during migration cycles, thus playing an important role in the export of nutrients (N, P, and Fe) from surface waters.[47][48][44]
Zooplankton feed in the euphotic zone and export nutrients via sinking fecal pellets, and vertical migration. Fish typically release nutrients at the same depth at which they feed. Excretion for marine mammals, tethered to the surface for respiration, is expected to be shallower in the water column than where they feed.[44]
Marine mammals provide important ecosystem services. On a global scale, they can influence climate, through fertilization events and the export of carbon from surface waters to the deep sea through sinking whale carcasses.[49] In coastal areas, whales retain nutrients locally, increasing ecosystem productivity and perhaps raising the carrying capacity for other marine consumers, including commercial fish species.[44] It has been estimated that, in terms of carbon sequestration, one whale is equivalent to thousands of trees.[50]
Lipid pump
The lipid pump is the sequestration of carbon from the ocean surface to deeper waters due to the presence of lipids (fats) in overwintering vertically migratory zooplankton. This carbon enters the deep ocean through the respiration and mortality of the migrating zooplankton. The lipid pump also entails a lipid shunt, where other nutrients like nitrogen and phosphorus that are consumed in excess are excreted back to the surface environment. This means that the carbon transported due to the lipid pump does not affect the essential nutrients at the ocean surface.[51] The contribution of the lipid pump to the sequestering of carbon in the deeper waters of the ocean can be substantial: the carbon transported below 1,000 metres (3,300 ft) by copepods of the genus Calanus in the Arctic Ocean almost equals that transported below the same depth annually by particulate organic carbon.[52]
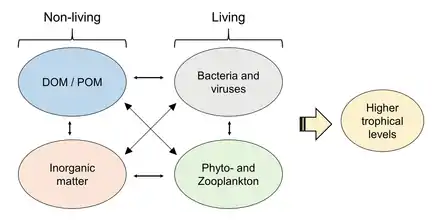
Microbial loop
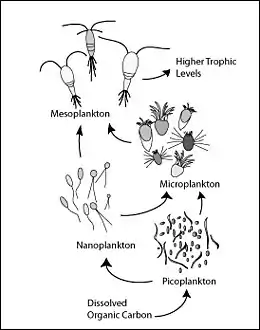
The microbial loop describes a trophic pathway in the marine microbial food web where dissolved organic carbon (DOC) is returned to higher trophic levels via its incorporation into bacterial biomass, and then coupled with the classic food chain formed by phytoplankton-zooplankton-nekton. The term microbial loop was coined by Farooq Azam, Tom Fenchel et al.[53] in 1983 to include the role played by bacteria in the carbon and nutrient cycles of the marine environment. In general, dissolved organic carbon (DOC) is introduced into the ocean environment from bacterial lysis, the leakage or exudation of fixed carbon from phytoplankton (e.g., mucilaginous exopolymer from diatoms), sudden cell senescence, sloppy feeding by zooplankton, the excretion of waste products by aquatic animals, or the breakdown or dissolution of organic particles from terrestrial plants and soils.[54] Bacteria in the microbial loop decompose this particulate detritus to utilize this energy-rich matter for growth. Since more than 95% of organic matter in marine ecosystems consists of polymeric, high molecular weight (HMW) compounds (e.g., protein, polysaccharides, lipids), only a small portion of total dissolved organic matter (DOM) is readily utilizable to most marine organisms at higher trophic levels. This means that dissolved organic carbon is not available directly to most marine organisms; marine bacteria introduce this organic carbon into the food web, resulting in additional energy becoming available to higher trophic levels.
Viral shunt
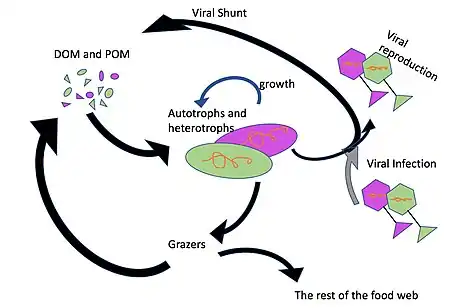
As much as 25% of the primary production from phytoplankton in the global oceans may be recycled within the microbial loop through viral shunting.[55] The viral shunt is a mechanism whereby marine viruses prevent microbial particulate organic matter (POM) from migrating up trophic levels by recycling them into dissolved organic matter (DOM), which can be readily taken up by microorganisms. The DOM recycled by the viral shunt pathway is comparable to the amount generated by the other main sources of marine DOM.[56] Viruses can easily infect microorganisms in the microbial loop due to their relative abundance compared to microbes.[57][58] Prokaryotic and eukaryotic mortality contribute to carbon nutrient recycling through cell lysis. There is evidence as well of nitrogen (specifically ammonium) regeneration. This nutrient recycling helps stimulates microbial growth.[59]
Bioluminescent shunt
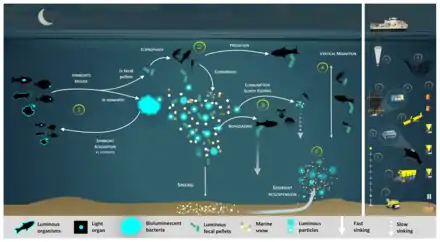
Luminous bacteria in light organ symbioses are successively acquired by host (squid, fish) from the seawater while they are juveniles, then regularly released into the ocean.[60]
In the diagram on the right, depending on the light organ position, luminous bacteria are released from their guts into fecal pellets or directly into the seawater (step 1). Motile luminous bacteria colonize organic matter sinking along the water column. Bioluminescent bacteria inseminating fecal pellets and particles influence zooplankton consumption rates. Such visual markers increase detection (“bait hypothesis”), attraction and finally predation by upper trophic levels (step 2). In the mesopelagic, zooplankton and their predators feed on sinking luminous particles and fecal pellets, which form either aggregates (repackaging) of faster sinking rates or fragment organic matter (due to sloppy feeding) with slower sinking rates (step 3).[60]
Filter feeders also aggregate sinking organic matter without particular visual detection and selection of luminous matter. Diel (and seasonal) vertical migrators feeding on luminous food metabolize and release glowing fecal pellets from the surface to the mesopelagic zone (step 4). This implies bioluminescent bacteria dispersion at large spatial scales, for zooplankton or even some fish actively swimming long distances. Luminous bacteria attached to particles sink down to the seafloor, and sediment can be resuspended by oceanographic physical conditions (step 5) and consumed by epi-benthic organisms. Instruments are (a) plankton net, (b) fish net, (c) Niskin water sampler, (d) bathyphotometer, (e) sediment traps, (f) autonomous underwater vehicles, (g) photomultiplier module, (h) astrophysics optical modules ANTARES and (i–j) remotely operated vehicles.[60]
Quantification

The geologic component of the carbon cycle operates slowly in comparison to the other parts of the global carbon cycle. It is one of the most important determinants of the amount of carbon in the atmosphere, and thus of global temperatures.[62]
As the biological pump plays an important role in the Earth's carbon cycle, significant effort is spent quantifying its strength. However, because they occur as a result of poorly constrained ecological interactions usually at depth, the processes that form the biological pump are difficult to measure. A common method is to estimate primary production fuelled by nitrate and ammonium as these nutrients have different sources that are related to the remineralisation of sinking material. From these it is possible to derive the so-called f-ratio, a proxy for the local strength of the biological pump. Applying the results of local studies to the global scale is complicated by the role the ocean's circulation plays in different ocean regions.[63]
Anthropogenic changes
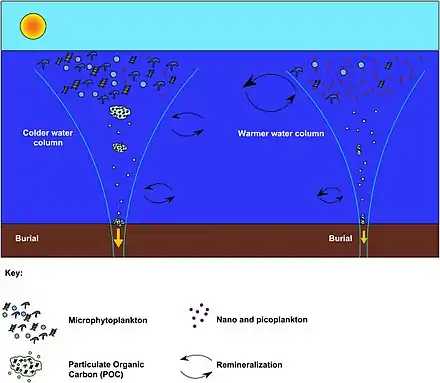
It was recently determined that coccolithophore concentrations in the North Atlantic have increased by an order of magnitude since the 1960s and an increase in absorbed CO2, as well as temperature, were modeled to be the most likely cause of this increase.[64]
Changes in land use, the combustion of fossil fuels, and the production of cement have led to an increase in CO2 concentration in the atmosphere. At present, about one third (approximately 2 Pg C y−1 = 2 × 1015 grams of carbon per year)[65][66] of anthropogenic emissions of CO2 are believed to be entering the ocean. However, the biological pump is not believed to play a significant role in the net uptake of CO2 by oceans. This is because the biological pump is primarily limited by the availability of light and nutrients, and not by carbon. This is in contrast to the situation on land, where elevated atmospheric concentrations of CO2 may increase primary production because land plants are able to improve their water-use efficiency (= decrease transpiration) when CO2 is easier to obtain.[67] However, there are still considerable uncertainties in the marine carbon cycle, and some research suggests that a link between elevated CO2 and marine primary production exists.[68]
[[File:Arctic carbon fluxes influenced by sea ice decline and permafrost thaw.webp|thumb|upright=2.0| {{center|Arctic carbon fluxes influenced by sea ice decline and permafrost thaw [69][70]]]
However, climate change may affect the biological pump in the future by warming and stratifying the surface ocean. It is believed that this could decrease the supply of nutrients to the euphotic zone, reducing primary production there. Also, changes in the ecological success of calcifying organisms caused by ocean acidification may affect the biological pump by altering the strength of the hard tissues pump.[71] This may then have a "knock-on" effect on the soft tissues pump because calcium carbonate acts to ballast sinking organic material.[72] In 2019, a study indicated that at current rates of seawater acidification, we could see Antarctic phytoplanktons smaller and less effective at storing carbon before the end of the century.[73]
The diagram on the right shows some possible effects of sea ice decline and permafrost thaw on Arctic carbon fluxes. On land, plants take up carbon while microorganisms in the soil produce methane and respire CO2. Lakes are net emitters of methane, and organic and inorganic carbon (dissolved and particulate) flow into the ocean through freshwater systems. In the ocean, methane can be released from thawing subsea permafrost, and CO2 is absorbed due to an undersaturation of CO2 in the water compared with the atmosphere. In addition, multiple fluxes are closely associated to sea ice. Current best estimates of atmospheric fluxes are given in Tg C year −1, where available. Note that the emission estimate for lakes is for the area North of ~50º N rather than the narrower definition of arctic tundra for the other terrestrial fluxes. When available, uncertainty ranges are shown in brackets. The arrows do not represent the size of each flux.[70][69]
Needed research
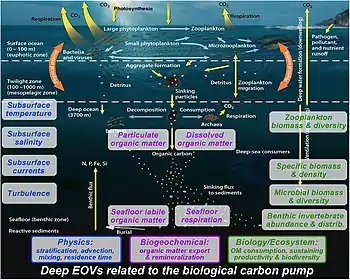
water column to properly understand the biological pump [74]
Multidisciplinary observations are still needed in the deep water column to properly understand the biological pump:[74]
- Physics: stratification affects particle sinking; understanding the origin of the particles and the residence time of the DIC from particle remineralization in the deep ocean requires measurement of advection and mixing.[74]
- Biogeochemistry: export/mixing down of particulate and dissolved organic matter from the surface layer determines labile organic matter arriving at the seafloor, which is either respired by seafloor biota or stored for longer times in the sediment.[74]
- Biology and ecosystems: zooplankton and microorganisms break down and remineralize sinking particles in the water column. Exported organic matter feeds all water column and benthic biota (zooplankton, benthic invertebrates, microbes) sustaining their biomass, density, and biodiversity.[74]
References
- Sigman DM & GH Haug. 2006. The biological pump in the past. In: Treatise on Geochemistry; vol. 6, (ed.). Pergamon Press, pp. 491-528
- Hain, M.P.; Sigman, D.M.; Haug, G.H. (2014). The Biological Pump in the Past (PDF). Treatise on Geochemistry, 2nd Edition. 8. pp. 485–517. doi:10.1016/B978-0-08-095975-7.00618-5. ISBN 9780080983004. Retrieved 2015-06-01.
- Boscolo-Galazzo, F.; Crichton, K.A.; Barker, S.; Pearson, P.N. (2018). "Temperature dependency of metabolic rates in the upper ocean: A positive feedback to global climate change?". Global and Planetary Change. 170: 201–212. Bibcode:2018GPC...170..201B. doi:10.1016/j.gloplacha.2018.08.017.
 Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License. - De La Rocha CL. 2006. The Biological Pump. In: Treatise on Geochemistry; vol. 6, (ed.). Pergamon Press, pp. 83-111
- Ducklow, H.W., Steinberg, D.K. and Buesseler, K.O. (2001) "Upper Ocean Carbon Export and the Biological Pump". Oceanography, 14(4): 50–58. doi:10.5670/oceanog.2001.06. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Monroy, P., Hernández-García, E., Rossi, V. and López, C. (2017) "Modeling the dynamical sinking of biogenic particles in oceanic flow". Nonlinear Processes in Geophysics, 24(2): 293–305. doi:10.5194/npg-24-293-2017. Material was copied from this source, which is available under a Creative Commons Attribution 3.0 International License.
- Simon, M., Grossart, H., Schweitzer, B. and Ploug, H. (2002) "Microbial ecology of organic aggregates in aquatic ecosystems". Aquatic microbial ecology, 28: 175–211. doi:10.3354/ame028175.
- Sigman, D.M.; Hain, M.P. (2012). "The Biological Productivity of the Ocean" (PDF). Nature Education Knowledge. 3 (6): 1–16. Retrieved 2015-06-01.
The value of NEP [Net Ecosystem Production] depends on the boundaries defined for the ecosystem. If one considers the sunlit surface ocean down to the 1% light level (the “euphotic zone”) over the course of an entire year, then NEP is equivalent to the particulate organic carbon sinking into the dark ocean interior plus the dissolved organic carbon being circulated out of the euphotic zone. In this case, NEP is also often referred to as “export production” (or “new production” (Dugdale & Goering 1967), as discussed below).
- Martiny, Adam C.; Pham, Chau T. A.; Primeau, Francois W.; Vrugt, Jasper A.; Moore, J. Keith; Levin, Simon A.; Lomas, Michael W. (April 2013). "Strong latitudinal patterns in the elemental ratios of marine plankton and organic matter". Nature Geoscience. 6 (4): 279–283. doi:10.1038/NGEO1757.
- Wilson, R. W.; Millero, F. J.; Taylor, J. R.; Walsh, P. J.; Christensen, V.; Jennings, S.; Grosell, M. (16 January 2009). "Contribution of Fish to the Marine Inorganic Carbon Cycle". Science. 323 (5912): 359–362. doi:10.1126/science.1157972.
- Pilson MEQ. 2012. An Introduction to the Chemistry of the Sea. Cambridge University Press, pp.
- Hain, M.P.; Sigman, D.M.; Haug, G.H. (2010). "Carbon dioxide effects of Antarctic stratification, North Atlantic Intermediate Water formation, and subantarctic nutrient drawdown during the last ice age: Diagnosis and synthesis in a geochemical box model". Global Biogeochemical Cycles. 24 (4): 1–19. Bibcode:2010GBioC..24.4023H. doi:10.1029/2010GB003790.
- H., Schlesinger, William (2013). Biogeochemistry : an analysis of global change. Bernhardt, Emily S. (3rd ed.). Waltham, Mass.: Academic Press. ISBN 9780123858740. OCLC 827935936.
- Falkowski, P.; Scholes, R. J.; Boyle, E.; Canadell, J.; Canfield, D.; Elser, J.; Gruber, N.; Hibbard, K.; Högberg, P. (2000-10-13). "The Global Carbon Cycle: A Test of Our Knowledge of Earth as a System". Science. 290 (5490): 291–296. Bibcode:2000Sci...290..291F. doi:10.1126/science.290.5490.291. ISSN 0036-8075. PMID 11030643.
- Raven, J. A.; P. G. Falkowski (1999). "Oceanic sinks for atmospheric CO2". Plant, Cell and Environment. 22 (6): 741–755. doi:10.1046/j.1365-3040.1999.00419.x.
- Raven, J. A.; Falkowski, P. G. (1999). "Oceanic sinks for atmospheric CO2". Plant, Cell and Environment. 22 (6): 741–755. doi:10.1046/j.1365-3040.1999.00419.x.
- Hain, M.P.; Sigman, D.M.; Haug, G.H (2014). "The Biological Pump in the Past". Treatise on Geochemistry. 8: 485-517. doi:10.1016/B978-0-08-095975-7.00618-5. ISBN 9780080983004.
- Rost, Bjorn; Reibessel, Ulf (2004). Coccolithophores and the biological pump: responses to environmental changes. Berlin, Heidelberg: Springer. ISBN 978-3-642-06016-8.
- Tsunogai, S.; Watanabe, S.; Sato, T. (1999). "Is there a "continental shelf pump" for the absorption of atmospheric CO2". Tellus B. 51 (3): 701–712. Bibcode:1999TellB..51..701T. doi:10.1034/j.1600-0889.1999.t01-2-00010.x.
- Wollast, R. (1998). Evaluation and comparison of the global carbon cycle in the coastal zone and in the open ocean, p. 213-252. In K. H. Brink and A. R. Robinson (eds.), The Global Coastal Ocean. John Wiley & Sons.
- Rippeth, T. P.; Scourse, J. D.; Uehara, K.; McKeown, S. (2008). "Impact of sea-level rise over the last deglacial transition on the strength of the continental shelf CO2 pump". Geophys. Res. Lett. 35 (24): L24604. Bibcode:2008GeoRL..3524604R. doi:10.1029/2008GL035880. S2CID 1049049.
- Heinrichs, Mara E.; Mori, Corinna; Dlugosch, Leon (2020). "Complex Interactions Between Aquatic Organisms and Their Chemical Environment Elucidated from Different Perspectives". YOUMARES 9 - the Oceans: Our Research, Our Future. pp. 279–297. doi:10.1007/978-3-030-20389-4_15. ISBN 978-3-030-20388-7. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Carlson CA (2002) [books.google.com/books?id=D6TMKZOgldAC&q=%22Production+and+removal+processes%22&pg=PA91 "Production and removal processes"]. In: Hansell DA, Carlson CA (eds) Biogeochemistry of marine dissolved organic matter. Academic Press, San Diego, pages 91–151. ISBN 9780123238412.
- Capelle, David W.; Kuzyk, Zou Zou A.; Papakyriakou, Tim; Guéguen, Céline; Miller, Lisa A.; MacDonald, Robie W. (2020). "Effect of terrestrial organic matter on ocean acidification and CO2 flux in an Arctic shelf sea". Progress in Oceanography. 185: 102319. Bibcode:2020PrOce.18502319C. doi:10.1016/j.pocean.2020.102319. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Cavan, E.L., Belcher, A., Atkinson, A., Hill, S.L., Kawaguchi, S., McCormack, S., Meyer, B., Nicol, S., Ratnarajah, L., Schmidt, K. and Steinberg, D.K. (2019) "The importance of Antarctic krill in biogeochemical cycles". Nature communications, 10(1): 1–13. doi:10.1038/s41467-019-12668-7. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Lampitt, R. S.; Noji, T.; von Bodungen, B. (1990). "What happens to zooplankton faecal pellets? Implications for material flux". Marine Biology. 104: 15–23. doi:10.1007/BF01313152. S2CID 86523326.
- Steinberg, Deborah; Sarah Goldthwait; Dennis Hansell (2002). "Zooplankton vertical migration and the active transport of dissolved organic and inorganic nitrogen in the Sargasso Sea". Deep-Sea Research Part I. 49 (8): 1445–1461. Bibcode:2002DSRI...49.1445S. CiteSeerX 10.1.1.391.7622. doi:10.1016/S0967-0637(02)00037-7. ISSN 0967-0637.
- Passow, U.; Carlson, C.A. (2012). "The biological pump in a high CO2 world" (PDF). Marine Ecology Progress Series. 470: 249–271. Bibcode:2012MEPS..470..249P. doi:10.3354/meps09985. Retrieved 25 February 2016.
- Robinson, J.; Popova, E.E.; Yool, A.; Srokosz, M.A.; Lampitt, R.S.; Blundell, J.R. (2014). "How deep is deep enough? Ocean iron fertilization and carbon sequestration in the Southern Ocean" (PDF). Geophys. Res. Lett. 41 (7): 2489–2495. Bibcode:2014GeoRL..41.2489R. doi:10.1002/2013GL058799.
- Møller, EF; Thor, P.; Nielsen, TG (2003). "Production of DOC by Calanus finmarchicus, C. Glacialis and C. Hyperboreus through sloppy feeding and leakage from fecal pellets". Marine Ecology Progress Series. 262: 185–191. Bibcode:2003MEPS..262..185M. doi:10.3354/meps262185.
- Saba, GK; Steinberg, DK; Bronk, DA (2009). "Effects of diet on release of dissolved organic and inorganic nutrients by the copepod Acartia tonsa". Marine Ecology Progress Series. 386: 147–161. Bibcode:2009MEPS..386..147S. doi:10.3354/meps08070.
- Steinberg, Deborah K.; Landry, Michael R. (2017). "Zooplankton and the Ocean Carbon Cycle". Annual Review of Marine Science. 9: 413–444. Bibcode:2017ARMS....9..413S. doi:10.1146/annurev-marine-010814-015924. PMID 27814033.
- Saba, Grace K.; Steinberg, Deborah K.; Bronk, Deborah A. (2011). "The relative importance of sloppy feeding, excretion, and fecal pellet leaching in the release of dissolved carbon and nitrogen by Acartia tonsa copepods". Journal of Experimental Marine Biology and Ecology. 404 (1–2): 47–56. doi:10.1016/j.jembe.2011.04.013.
- Thor, P.; Dam, HG; Rogers, DR (2003). "Fate of organic carbon released from decomposing copepod fecal pellets in relation to bacterial production and ectoenzymatic activity". Aquatic Microbial Ecology. 33: 279–288. doi:10.3354/ame033279.
- Hansell, Dennis A.; Carlson, Craig A. (2 October 2014). Biogeochemistry of Marine Dissolved Organic Matter. ISBN 9780124071537.
- Moller, E. F. (2004). "Sloppy feeding in marine copepods: Prey-size-dependent production of dissolved organic carbon". Journal of Plankton Research. 27: 27–35. doi:10.1093/plankt/fbh147.
- Møller, Eva Friis (2007). "Production of dissolved organic carbon by sloppy feeding in the copepods Acartia tonsa, Centropages typicus, and Temora longicornis". Limnology and Oceanography. 52 (1): 79–84. Bibcode:2007LimOc..52...79M. doi:10.4319/lo.2007.52.1.0079.
- Wiebe, P.H; L.P. Madin; L.R. Haury; G.R. Harbison; L.M. Philbin (1979). "Diel Vertical Migration by Salpa aspera and its potential for large-scale particulate organic matter transport to the deep-sea". Marine Biology. 53 (3): 249–255. doi:10.1007/BF00952433.
- Hays, Graeme C.; Doyle, Thomas K.; Houghton, Jonathan D.R. (2018). "A Paradigm Shift in the Trophic Importance of Jellyfish?". Trends in Ecology & Evolution. 33 (11): 874–884. doi:10.1016/j.tree.2018.09.001. PMID 30245075.
- Lebrato, Mario; Pitt, Kylie A.; Sweetman, Andrew K.; Jones, Daniel O. B.; Cartes, Joan E.; Oschlies, Andreas; Condon, Robert H.; Molinero, Juan Carlos & Adler, Laetitia (2012). "Jelly-falls historic and recent observations: a review to drive future research directions". Hydrobiologia. 690 (1): 227–245. doi:10.1007/s10750-012-1046-8.
- Lebrato, M. & Jones, D. O. B. (2009). "Mass deposition event of Pyrosoma atlanticum carcasses off Ivory Coast (West Africa)" (PDF). Limnology and Oceanography. 54 (4): 1197–1209. Bibcode:2009LimOc..54.1197L. doi:10.4319/lo.2009.54.4.1197.
- Sweetman, Andrew K. & Chapman, Annelise (2011). "First observations of jelly-falls at the seafloor in a deep-sea fjord". Deep Sea Research Part I: Oceanographic Research Papers. 58 (12): 1206–1211. Bibcode:2011DSRI...58.1206S. doi:10.1016/j.dsr.2011.08.006.
- Burd, Adrian. "Towards a transformative understanding of the ocean's biological pump: Priorities for future research-Report on the NSF Biology of the Biological Pump Workshop" (PDF). OCB: Ocean Carbon & Biogeochemistry. Retrieved 30 October 2016.
- Roman, Joe; McCarthy, James J. (2010). "The Whale Pump: Marine Mammals Enhance Primary Productivity in a Coastal Basin". PLOS ONE. 5 (10): e13255. Bibcode:2010PLoSO...513255R. doi:10.1371/journal.pone.0013255. PMC 2952594. PMID 20949007. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Brown, Joshua E. (12 Oct 2010). "Whale poop pumps up ocean health". Science Daily. Retrieved 18 August 2014.
- Longhurst, Alan R.; Glen Harrison, W. (1989). "The biological pump: Profiles of plankton production and consumption in the upper ocean". Progress in Oceanography. 22: 47–123. doi:10.1016/0079-6611(89)90010-4.
- Hutchins, David A.; Wang, Wen-Xiong; Fisher, Nicholas S. (1995). "Copepod grazing and the biogeochemical fate of diatom iron". Limnology and Oceanography. 40 (5): 989–994. doi:10.4319/lo.1995.40.5.0989.
- Steinberg, Deborah K.; Goldthwait, Sarah A.; Hansell, Dennis A. (2002). "Zooplankton vertical migration and the active transport of dissolved organic and inorganic nitrogen in the Sargasso Sea". Deep Sea Research Part I: Oceanographic Research Papers. 49 (8): 1445–1461. doi:10.1016/S0967-0637(02)00037-7.
- Pershing, Andrew J.; Christensen, Line B.; Record, Nicholas R.; Sherwood, Graham D.; Stetson, Peter B. (2010). "The Impact of Whaling on the Ocean Carbon Cycle: Why Bigger Was Better". PLOS ONE. 5 (8): e12444. Bibcode:2010PLoSO...512444P. doi:10.1371/journal.pone.0012444. PMC 2928761. PMID 20865156.
- "Nature's Solution to Climate Change – IMF F&D". International Monetary Fund. Retrieved 2 November 2019.
- Jónasdóttir, Sigrún Huld; Visser, André W.; Richardson, Katherine; Heath, Michael R. (2015). "Seasonal copepod lipid pump promotes carbon sequestration in the deep North Atlantic" (PDF). Proceedings of the National Academy of Sciences. 112 (39): 12122–12126. doi:10.1073/pnas.1512110112. ISSN 0027-8424.
- Steinberg, Deborah K.; Landry, Michael R. (2017). "Zooplankton and the ocean carbon cycle". Annual Review of Marine Science. 9 (1): 413–444. doi:10.1146/annurev-marine-010814-015924. ISSN 1941-1405. PMID 27814033.
- Azam, Farooq; Fenchel, Tom; Field, J.G.; Gray, J.S.; Meyer-Reil, L.A.; Thingstad, F. (1983). "The Ecological Role of Water-Column Microbes in the Sea". Marine Ecology Progress Series. 10: 257–263. doi:10.3354/meps010257.
- Van den Meersche, Karel; Middelburg, Jack J.; Soetaert, Karline; van Rijswijk, Pieter; Boschker, Henricus T. S.; Heip, Carlo H. R. (2004). "Carbon-nitrogen coupling and algal-bacterial interactions during an experimental bloom: Modeling a13C tracer experiment". Limnology and Oceanography. 49 (3): 862–878. doi:10.4319/lo.2004.49.3.0862. ISSN 0024-3590.
- Wilhelm, Steven W.; Suttle, Curtis A. (1999). "Viruses and nutrient cycles in the sea: viruses play critical roles in the structure and function of aquatic food webs". BioScience. 49 (10): 781–788. doi:10.2307/1313569. JSTOR 1313569.
- Robinson, Carol, and Nagappa Ramaiah. "Microbial heterotrophic metabolic rates constrain the microbial carbon pump." The American Association for the Advancement of Science, 2011.
- Fuhrman, Jed A. (1999). "Marine viruses and their biogeochemical and ecological effects". Nature. 399 (6736): 541–548. Bibcode:1999Natur.399..541F. doi:10.1038/21119. ISSN 0028-0836. PMID 10376593.
- Wigington, Charles H.; Sonderegger, Derek; Brussaard, Corina P. D.; Buchan, Alison; Finke, Jan F.; Fuhrman, Jed A.; Lennon, Jay T.; Middelboe, Mathias; Suttle, Curtis A.; Stock, Charles; Wilson, William H. (March 2016). "Re-examination of the relationship between marine virus and microbial cell abundances". Nature Microbiology. 1 (3): 15024. doi:10.1038/nmicrobiol.2015.24. ISSN 2058-5276.
- Tsai, An-Yi, Gwo-Ching Gong, and Yu-Wen Huang. "Importance of the Viral Shunt in Nitrogen Cycling in Synechococcus Spp. Growth in Subtropical Western Pacific Coastal Waters." Terrestrial, Atmospheric & Oceanic Sciences25.6 (2014).
- Tanet, Lisa; Martini, Séverine; Casalot, Laurie; Tamburini, Christian (2020). "Reviews and syntheses: Bacterial bioluminescence – ecology and impact in the biological carbon pump". Biogeosciences. 17 (14): 3757–3778. Bibcode:2020BGeo...17.3757T. doi:10.5194/bg-17-3757-2020. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Kayler, Z., Janowiak, M., Swanston, C. (2017). "The Global Carbon Cycle". Considering Forest and Grassland Carbon in Land Management. General Technical Report WTO-GTR-95. United States Department of Agriculture, Forest Service. pp. 3–9.CS1 maint: multiple names: authors list (link)
- NASA (16 June 2011). "The Slow Carbon Cycle". Archived from the original on 16 June 2012. Retrieved 24 June 2012.
- Marinov, I.; Gnanadesikan, A.; Toggweiler, J. R.; Sarmiento, J. L. (2006). "The Southern Ocean biogeochemical divide". Nature. 441 (7096): 964–967. Bibcode:2006Natur.441..964M. doi:10.1038/nature04883. PMID 16791191.
- Rivero-Calle, Sara; Gnanadesikan, Anand; Castillo, Carlos E. Del; Balch, William M. & Guikema, Seth D. (2015). "Multidecadal increase in North Atlantic coccolithophores and the potential role of rising CO2". Science. 350 (6267): 1533–1537. Bibcode:2015Sci...350.1533R. doi:10.1126/science.aaa8026. PMID 26612836.
- Takahashi, Taro; Sutherland, Stewart C.; Sweeney, Colm; Poisson, Alain; Metzl, Nicolas; Tilbrook, Bronte; Bates, Nicolas; Wanninkhof, Rik; Feely, Richard A.; Sabine, Christopher; Olafsson, Jon; Nojiri, Yukihiro (2002). "Global sea–air CO2 flux based on climatological surface ocean pCO2, and seasonal biological and temperature effects". Deep Sea Research Part II: Topical Studies in Oceanography. 49 (9–10): 1601–1622. Bibcode:2002DSRII..49.1601T. doi:10.1016/S0967-0645(02)00003-6.
- Orr, J. C., E. Maier-Reimer, U. Mikolajewicz, P. Monfray, J. L. Sarmiento, J. R. Toggweiler, N. K. Taylor, J. Palmer, N. Gruber, C. L. Sabine, C. Le Quéré, R. M. Key and J. Boutin (2001). Estimates of anthropogenic carbon uptake from four three-dimensional global ocean models. Global Biogeochem. Cycles 15, 43–60.
- Cox, P. M., Betts, R. A., Jones, C. D., Spall, S. A. and Totterdell, I. J. (2000). Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model. Nature, 408, 184–187.
- Riebesell, U., Schulz, K.G., Bellerby, R.G.J., Botros, M., Fritsche, P., Meyerhöfer, M., Neill, C., Nondal, G., Oschlies, A., Wohlers, J. and Zöllner, E. (2007). Enhanced biological carbon consumption in a high CO2 ocean. Nature 450, 545–548.
- Parmentier, Frans-Jan W.; Christensen, Torben R.; Rysgaard, Søren; Bendtsen, Jørgen; Glud, Ronnie N.; Else, Brent; Van Huissteden, Jacobus; Sachs, Torsten; Vonk, Jorien E.; Sejr, Mikael K. (2017). "A synthesis of the arctic terrestrial and marine carbon cycles under pressure from a dwindling cryosphere". Ambio. 46 (Suppl 1): 53–69. doi:10.1007/s13280-016-0872-8. PMC 5258664. PMID 28116680. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.
- Parmentier, Frans-Jan W.; Christensen, Torben R.; Sørensen, Lise Lotte; Rysgaard, Søren; McGuire, A. David; Miller, Paul A.; Walker, Donald A. (2013). "The impact of lower sea-ice extent on Arctic greenhouse-gas exchange". Nature Climate Change. 3 (3): 195–202. doi:10.1038/nclimate1784.
- Orr, James C.; Fabry, Victoria J.; Aumont, Olivier; Bopp, Laurent; Doney, Scott C.; Feely, Richard A.; Gnanadesikan, Anand; Gruber, Nicolas; Ishida, Akio; et al. (2005). "Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms" (PDF). Nature. 437 (7059): 681–686. Bibcode:2005Natur.437..681O. doi:10.1038/nature04095. PMID 16193043.
- Armstrong, Robert A.; Lee, Cindy; Hedges, John I.; Honjo, Susumu & Wakeham, Stuart G. (2001). "A new, mechanistic model for organic carbon fluxes in the ocean based on the quantitative association of POC with ballast minerals". Deep Sea Research Part II: Topical Studies in Oceanography. 49 (1–3): 219–236. Bibcode:2001DSRII..49..219A. doi:10.1016/s0967-0645(01)00101-1.
- Petrou, Katherina; Nielsen, Daniel (2019-08-27). "Acid oceans are shrinking plankton, fueling faster climate change". phys.org. Retrieved 2019-09-07.
- Levin, L.A., Bett, B.J., Gates, A.R., Heimbach, P., Howe, B.M., Janssen, F., McCurdy, A., Ruhl, H.A., Snelgrove, P., Stocks, K.I., Bailey, D. and 27 others (2019) "Global observing needs in the deep ocean". Frontiers in Marine Science, 6: 241. doi:10.3389/fmars.2019.00241. Material was copied from this source, which is available under a Creative Commons Attribution 4.0 International License.