Collagen
Collagen (/ˈkɒlədʒɪn/) is the main structural protein in the extracellular matrix found in the body's various connective tissues. As the main component of connective tissue, it is the most abundant protein in mammals,[1] making up from 25% to 35% of the whole-body protein content. Collagen consists of amino acids bound together to form a triple helix of elongated fibril[2] known as a collagen helix. It is mostly found in connective tissue such as cartilage, bones, tendons, ligaments, and skin.
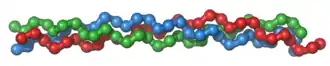
Depending upon the degree of mineralization, collagen tissues may be rigid (bone), compliant (tendon), or have a gradient from rigid to compliant (cartilage). Collagen is also abundant in corneas, blood vessels, the gut, intervertebral discs, and the dentin in teeth.[3] In muscle tissue, it serves as a major component of the endomysium. Collagen constitutes one to two percent of muscle tissue and accounts for 6% of the weight of strong, tendinous, muscles.[4] The fibroblast is the most common cell that creates collagen. Gelatin, which is used in food and industry, is collagen that has been irreversibly hydrolyzed.[5] Collagen has many medical uses in treating complications of the bones and skin.
The name collagen comes from the Greek κόλλα (kólla), meaning "glue", and suffix -γέν, -gen, denoting "producing".[6][7] This refers to the compound's early use in the process of creating glue from boiling the skin and tendons of horses and other animals.
Types
Over 90% of the collagen in the human body is type I collagen.[8] However, as of 2011, 30 types of collagen have been identified, described, and divided into several groups according to the structure they form.[9] All of the types contain at least one triple helix.[9] The number of types shows collagen's diverse functionality.[10]
- Fibrillar (Type I, II, III, V, XI)
- Non-fibrillar
- FACIT (Fibril Associated Collagens with Interrupted Triple Helices) (Type IX, XII, XIV, XIX, XXI)
- Short chain (Type VIII, X)
- Basement membrane (Type IV)
- Multiplexin (Multiple Triple Helix domains with Interruptions) (Type XV, XVIII)
- MACIT (Membrane Associated Collagens with Interrupted Triple Helices) (Type XIII, XVII)
- Microfibril forming (Type VI)
- Anchoring fibrils (Type VII)
The five most common types are:
- Type I: skin, tendon, vasculature, organs, bone (main component of the organic part of bone)
- Type II: cartilage (main collagenous component of cartilage)
- Type III: reticulate (main component of reticular fibers), commonly found alongside type I
- Type IV: forms basal lamina, the epithelium-secreted layer of the basement membrane
- Type V: cell surfaces, hair, and placenta
Medical uses
Cardiac applications
The collagenous cardiac skeleton which includes the four heart valve rings, is histologically, elastically and uniquely bound to cardiac muscle. The cardiac skeleton also includes the separating septa of the heart chambers – the interventricular septum and the atrioventricular septum. Collagen contribution to the measure of cardiac performance summarily represents a continuous torsional force opposed to the fluid mechanics of blood pressure emitted from the heart. The collagenous structure that divides the upper chambers of the heart from the lower chambers is an impermeable membrane that excludes both blood and electrical impulses through typical physiological means. With support from collagen, atrial fibrillation never deteriorates to ventricular fibrillation. Collagen is layered in variable densities with smooth muscle mass. The mass, distribution, age and density of collagen all contribute to the compliance required to move blood back and forth. Individual cardiac valvular leaflets are folded into shape by specialized collagen under variable pressure. Gradual calcium deposition within collagen occurs as a natural function of aging. Calcified points within collagen matrices show contrast in a moving display of blood and muscle, enabling methods of cardiac imaging technology to arrive at ratios essentially stating blood in (cardiac input) and blood out (cardiac output). Pathology of the collagen underpinning of the heart is understood within the category of connective tissue disease.
Cosmetic surgery
Collagen has been widely used in cosmetic surgery, as a healing aid for burn patients for reconstruction of bone and a wide variety of dental, orthopedic, and surgical purposes. Both human and bovine collagen is widely used as dermal fillers for treatment of wrinkles and skin aging.[11] Some points of interest are:
- When used cosmetically, there is a chance of allergic reactions causing prolonged redness; however, this can be virtually eliminated by simple and inconspicuous patch testing prior to cosmetic use.
- Most medical collagen is derived from young beef cattle (bovine) from certified BSE-free animals. Most manufacturers use donor animals from either "closed herds", or from countries which have never had a reported case of BSE such as Australia, Brazil, and New Zealand.
Bone grafts
As the skeleton forms the structure of the body, it is vital that it maintains its strength, even after breaks and injuries. Collagen is used in bone grafting as it has a triple helical structure, making it a very strong molecule. It is ideal for use in bones, as it does not compromise the structural integrity of the skeleton. The triple helical structure of collagen prevents it from being broken down by enzymes, it enables adhesiveness of cells and it is important for the proper assembly of the extracellular matrix.[12]
Tissue regeneration
Collagen scaffolds are used in tissue regeneration, whether in sponges,[13] thin sheets,[14] gels,[15] or fibers.[16] Collagen has favorable properties for tissue regeneration, such as pore structure, permeability, hydrophilicity, and stability in vivo. Collagen scaffolds also support deposition of cells, such as osteoblasts and fibroblasts, and once inserted, facilitate growth to proceed normally.[17]
Reconstructive surgical uses
Collagens are widely employed in the construction of artificial skin substitutes used in the management of severe burns and wounds.[18][19] These collagens may be derived from bovine, equine, porcine, or even human sources; and are sometimes used in combination with silicones, glycosaminoglycans, fibroblasts, growth factors and other substances.
Wound healing
Collagen is one of the body's key natural resources and a component of skin tissue that can benefit all stages of wound healing.[20] When collagen is made available to the wound bed, closure can occur. Wound deterioration, followed sometimes by procedures such as amputation, can thus be avoided.
Collagen is a natural product and is thus used as a natural wound dressing and has properties that artificial wound dressings do not have. It is resistant against bacteria, which is of vital importance in a wound dressing. It helps to keep the wound sterile, because of its natural ability to fight infection. When collagen is used as a burn dressing, healthy granulation tissue is able to form very quickly over the burn, helping it to heal rapidly.[21]
Throughout the 4 phases of wound healing, collagen performs the following functions in wound healing:
- Guiding function: Collagen fibers serve to guide fibroblasts. Fibroblasts migrate along a connective tissue matrix.
- Chemotactic properties: The large surface area available on collagen fibers can attract fibrogenic cells which help in healing.
- Nucleation: Collagen, in the presence of certain neutral salt molecules can act as a nucleating agent causing formation of fibrillar structures. A collagen wound dressing might serve as a guide for orienting new collagen deposition and capillary growth.
- Hemostatic properties: Blood platelets interact with the collagen to make a hemostatic plug.
Basic research
Collagen is used in laboratory studies for cell culture, studying cell behavior and cellular interactions with the extracellular environment.[22]
Chemistry
The collagen protein is composed of a triple helix, which generally consists of two identical chains (α1) and an additional chain that differs slightly in its chemical composition (α2).[23] The amino acid composition of collagen is atypical for proteins, particularly with respect to its high hydroxyproline content. The most common motifs in the amino acid sequence of collagen are glycine-proline-X and glycine-X-hydroxyproline, where X is any amino acid other than glycine, proline or hydroxyproline. The average amino acid composition for fish and mammal skin is given.[23]
| Amino acid | Abundance in mammal skin (residues/1000) | Abundance in fish skin (residues/1000) |
|---|---|---|
| Glycine | 329 | 339 |
| Proline | 126 | 108 |
| Alanine | 109 | 114 |
| Hydroxyproline | 95 | 67 |
| Glutamic acid | 74 | 76 |
| Arginine | 49 | 52 |
| Aspartic acid | 47 | 47 |
| Serine | 36 | 46 |
| Lysine | 29 | 26 |
| Leucine | 24 | 23 |
| Valine | 22 | 21 |
| Threonine | 19 | 26 |
| Phenylalanine | 13 | 14 |
| Isoleucine | 11 | 11 |
| Hydroxylysine | 6 | 8 |
| Methionine | 6 | 13 |
| Histidine | 5 | 7 |
| Tyrosine | 3 | 3 |
| Cysteine | 1 | 1 |
| Tryptophan | 0 | 0 |
Synthesis
First, a three-dimensional stranded structure is assembled, with the amino acids glycine and proline as its principal components. This is not yet collagen but its precursor, procollagen. Procollagen is then modified by the addition of hydroxyl groups to the amino acids proline and lysine. This step is important for later glycosylation and the formation of the triple helix structure of collagen. Because the hydroxylase enzymes that perform these reactions require vitamin C as a cofactor, a long-term deficiency in this vitamin results in impaired collagen synthesis and scurvy.[24] These hydroxylation reactions are catalyzed by two different enzymes: prolyl-4-hydroxylase[25] and lysyl-hydroxylase. The reaction consumes one ascorbate molecule per hydroxylation. [26] The synthesis of collagen occurs inside and outside of the cell. The formation of collagen which results in fibrillary collagen (most common form) is discussed here. Meshwork collagen, which is often involved in the formation of filtration systems, is the other form of collagen. All types of collagens are triple helices, and the differences lie in the make-up of the alpha peptides created in step 2.
- Transcription of mRNA: About 44 genes are associated with collagen formation, each coding for a specific mRNA sequence, and typically have the "COL" prefix. The beginning of collagen synthesis begins with turning on genes which are associated with the formation of a particular alpha peptide (typically alpha 1, 2 or 3).
- Pre-pro-peptide formation: Once the final mRNA exits from the cell nucleus and enters into the cytoplasm, it links with the ribosomal subunits and the process of translation occurs. The early/first part of the new peptide is known as the signal sequence. The signal sequence on the N-terminal of the peptide is recognized by a signal recognition particle on the endoplasmic reticulum, which will be responsible for directing the pre-pro-peptide into the endoplasmic reticulum. Therefore, once the synthesis of new peptide is finished, it goes directly into the endoplasmic reticulum for post-translational processing. It is now known as preprocollagen.
- Pre-pro-peptide to pro-collagen: Three modifications of the pre-pro-peptide occur leading to the formation of the alpha peptide:
- The signal peptide on the N-terminal is removed, and the molecule is now known as propeptide (not procollagen).
- Hydroxylation of lysines and prolines on propeptide by the enzymes 'prolyl hydroxylase' and 'lysyl hydroxylase' (to produce hydroxyproline and hydroxylysine) occurs to aid cross-linking of the alpha peptides. This enzymatic step requires vitamin C as a cofactor. In scurvy, the lack of hydroxylation of prolines and lysines causes a looser triple helix (which is formed by three alpha peptides).
- Glycosylation occurs by adding either glucose or galactose monomers onto the hydroxyl groups that were placed onto lysines, but not on prolines.
- Once these modifications have taken place, three of the hydroxylated and glycosylated propeptides twist into a triple helix forming procollagen. Procollagen still has unwound ends, which will be later trimmed. At this point, the procollagen is packaged into a transfer vesicle destined for the Golgi apparatus.
- Golgi apparatus modification: In the Golgi apparatus, the procollagen goes through one last post-translational modification before being secreted out of the cell. In this step, oligosaccharides (not monosaccharides as in step 3) are added, and then the procollagen is packaged into a secretory vesicle destined for the extracellular space.
- Formation of tropocollagen: Once outside the cell, membrane bound enzymes known as collagen peptidases, remove the "loose ends" of the procollagen molecule. What is left is known as tropocollagen. Defects in this step produce one of the many collagenopathies known as Ehlers-Danlos syndrome. This step is absent when synthesizing type III, a type of fibrilar collagen.
- Formation of the collagen fibril: lysyl oxidase, an extracellular copper-dependent enzyme, produces the final step in the collagen synthesis pathway. This enzyme acts on lysines and hydroxylysines producing aldehyde groups, which will eventually undergo covalent bonding between tropocollagen molecules. This polymer of tropocollogen is known as a collagen fibril.
.svg.png.webp)
Amino acids
Collagen has an unusual amino acid composition and sequence:
- Glycine is found at almost every third residue.
- Proline makes up about 17% of collagen.
- Collagen contains two uncommon derivative amino acids not directly inserted during translation. These amino acids are found at specific locations relative to glycine and are modified post-translationally by different enzymes, both of which require vitamin C as a cofactor.
- Hydroxyproline derived from proline
- Hydroxylysine derived from lysine – depending on the type of collagen, varying numbers of hydroxylysines are glycosylated (mostly having disaccharides attached).
Cortisol stimulates degradation of (skin) collagen into amino acids.[27]
Collagen I formation
Most collagen forms in a similar manner, but the following process is typical for type I:
- Inside the cell
- Two types of alpha chains – alpha-1 and alpha 2, are formed during translation on ribosomes along the rough endoplasmic reticulum (RER). These peptide chains known as preprocollagen, have registration peptides on each end and a signal peptide.[28]
- Polypeptide chains are released into the lumen of the RER.
- Signal peptides are cleaved inside the RER and the chains are now known as pro-alpha chains.
- Hydroxylation of lysine and proline amino acids occurs inside the lumen. This process is dependent on and consumes ascorbic acid (vitamin C) as a cofactor.
- Glycosylation of specific hydroxylysine residues occurs.
- Triple alpha helical structure is formed inside the endoplasmic reticulum from two alpha-1 chains and one alpha-2 chain.
- Procollagen is shipped to the Golgi apparatus, where it is packaged and secreted by exocytosis.
- Outside the cell
- Registration peptides are cleaved and tropocollagen is formed by procollagen peptidase.
- Multiple tropocollagen molecules form collagen fibrils, via covalent cross-linking (aldol reaction) by lysyl oxidase which links hydroxylysine and lysine residues. Multiple collagen fibrils form into collagen fibers.
- Collagen may be attached to cell membranes via several types of protein, including fibronectin, laminin, fibulin and integrin.
Synthetic pathogenesis
Vitamin C deficiency causes scurvy, a serious and painful disease in which defective collagen prevents the formation of strong connective tissue. Gums deteriorate and bleed, with loss of teeth; skin discolors, and wounds do not heal. Prior to the 18th century, this condition was notorious among long-duration military, particularly naval, expeditions during which participants were deprived of foods containing vitamin C.
An autoimmune disease such as lupus erythematosus or rheumatoid arthritis[29] may attack healthy collagen fibers.
Many bacteria and viruses secrete virulence factors, such as the enzyme collagenase, which destroys collagen or interferes with its production.
Molecular structure
A single collagen molecule, tropocollagen, is used to make up larger collagen aggregates, such as fibrils. It is approximately 300 nm long and 1.5 nm in diameter, and it is made up of three polypeptide strands (called alpha peptides, see step 2), each of which has the conformation of a left-handed helix – this should not be confused with the right-handed alpha helix. These three left-handed helices are twisted together into a right-handed triple helix or "super helix", a cooperative quaternary structure stabilized by many hydrogen bonds. With type I collagen and possibly all fibrillar collagens, if not all collagens, each triple-helix associates into a right-handed super-super-coil referred to as the collagen microfibril. Each microfibril is interdigitated with its neighboring microfibrils to a degree that might suggest they are individually unstable, although within collagen fibrils, they are so well ordered as to be crystalline.
.png.webp)
A distinctive feature of collagen is the regular arrangement of amino acids in each of the three chains of these collagen subunits. The sequence often follows the pattern Gly-Pro-X or Gly-X-Hyp, where X may be any of various other amino acid residues.[23] Proline or hydroxyproline constitute about 1/6 of the total sequence. With glycine accounting for the 1/3 of the sequence, this means approximately half of the collagen sequence is not glycine, proline or hydroxyproline, a fact often missed due to the distraction of the unusual GX1X2 character of collagen alpha-peptides. The high glycine content of collagen is important with respect to stabilization of the collagen helix as this allows the very close association of the collagen fibers within the molecule, facilitating hydrogen bonding and the formation of intermolecular cross-links.[23] This kind of regular repetition and high glycine content is found in only a few other fibrous proteins, such as silk fibroin.
Collagen is not only a structural protein. Due to its key role in the determination of cell phenotype, cell adhesion, tissue regulation, and infrastructure, many sections of its non-proline-rich regions have cell or matrix association/regulation roles. The relatively high content of proline and hydroxyproline rings, with their geometrically constrained carboxyl and (secondary) amino groups, along with the rich abundance of glycine, accounts for the tendency of the individual polypeptide strands to form left-handed helices spontaneously, without any intrachain hydrogen bonding.
Because glycine is the smallest amino acid with no side chain, it plays a unique role in fibrous structural proteins. In collagen, Gly is required at every third position because the assembly of the triple helix puts this residue at the interior (axis) of the helix, where there is no space for a larger side group than glycine's single hydrogen atom. For the same reason, the rings of the Pro and Hyp must point outward. These two amino acids help stabilize the triple helix—Hyp even more so than Pro; a lower concentration of them is required in animals such as fish, whose body temperatures are lower than most warm-blooded animals. Lower proline and hydroxyproline contents are characteristic of cold-water, but not warm-water fish; the latter tend to have similar proline and hydroxyproline contents to mammals.[23] The lower proline and hydroxproline contents of cold-water fish and other poikilotherm animals leads to their collagen having a lower thermal stability than mammalian collagen.[23] This lower thermal stability means that gelatin derived from fish collagen is not suitable for many food and industrial applications.
The tropocollagen subunits spontaneously self-assemble, with regularly staggered ends, into even larger arrays in the extracellular spaces of tissues.[30][31] Additional assembly of fibrils is guided by fibroblasts, which deposit fully formed fibrils from fibripositors. In the fibrillar collagens, molecules are staggered to adjacent molecules by about 67 nm (a unit that is referred to as ‘D’ and changes depending upon the hydration state of the aggregate). In each D-period repeat of the microfibril, there is a part containing five molecules in cross-section, called the "overlap", and a part containing only four molecules, called the "gap".[32] These overlap and gap regions are retained as microfibrils assemble into fibrils, and are thus viewable using electron microscopy. The triple helical tropocollagens in the microfibrils are arranged in a quasihexagonal packing pattern.[32][33]

There is some covalent crosslinking within the triple helices, and a variable amount of covalent crosslinking between tropocollagen helices forming well organized aggregates (such as fibrils).[34] Larger fibrillar bundles are formed with the aid of several different classes of proteins (including different collagen types), glycoproteins, and proteoglycans to form the different types of mature tissues from alternate combinations of the same key players.[31] Collagen's insolubility was a barrier to the study of monomeric collagen until it was found that tropocollagen from young animals can be extracted because it is not yet fully crosslinked. However, advances in microscopy techniques (i.e. electron microscopy (EM) and atomic force microscopy (AFM)) and X-ray diffraction have enabled researchers to obtain increasingly detailed images of collagen structure in situ.[35] These later advances are particularly important to better understanding the way in which collagen structure affects cell–cell and cell–matrix communication and how tissues are constructed in growth and repair and changed in development and disease.[36][37] For example, using AFM–based nanoindentation it has been shown that a single collagen fibril is a heterogeneous material along its axial direction with significantly different mechanical properties in its gap and overlap regions, correlating with its different molecular organizations in these two regions.[38]
Collagen fibrils/aggregates are arranged in different combinations and concentrations in various tissues to provide varying tissue properties. In bone, entire collagen triple helices lie in a parallel, staggered array. 40 nm gaps between the ends of the tropocollagen subunits (approximately equal to the gap region) probably serve as nucleation sites for the deposition of long, hard, fine crystals of the mineral component, which is hydroxylapatite (approximately) Ca10(OH)2(PO4)6.[39] Type I collagen gives bone its tensile strength.
Associated disorders
Collagen-related diseases most commonly arise from genetic defects or nutritional deficiencies that affect the biosynthesis, assembly, postranslational modification, secretion, or other processes involved in normal collagen production.
| Type | Notes | Gene(s) | Disorders |
| I | This is the most abundant collagen of the human body. It is present in Scar tissue, the end product when tissue heals by repair. It is found in tendons, skin, artery walls, cornea, the endomysium surrounding muscle fibers, fibrocartilage, and the organic part of bones and teeth. | COL1A1, COL1A2 | Osteogenesis imperfecta, Ehlers–Danlos syndrome, infantile cortical hyperostosis a.k.a. Caffey's disease |
| II | Hyaline cartilage, makes up 50% of all cartilage protein. Vitreous humour of the eye. | COL2A1 | Collagenopathy, types II and XI |
| III | This is the collagen of granulation tissue and is produced quickly by young fibroblasts before the tougher type I collagen is synthesized. Reticular fiber. Also found in artery walls, skin, intestines and the uterus | COL3A1 | Ehlers–Danlos syndrome, Dupuytren's contracture |
| IV | Basal lamina; eye lens. Also serves as part of the filtration system in capillaries and the glomeruli of nephron in the kidney. | COL4A1, COL4A2, COL4A3, COL4A4, COL4A5, COL4A6 | Alport syndrome, Goodpasture's syndrome |
| V | Most interstitial tissue, assoc. with type I, associated with placenta | COL5A1, COL5A2, COL5A3 | Ehlers–Danlos syndrome (classical) |
| VI | Most interstitial tissue, assoc. with type I | COL6A1, COL6A2, COL6A3, COL6A5 | Ulrich myopathy, Bethlem myopathy, atopic dermatitis[40] |
| VII | Forms anchoring fibrils in dermoepidermal junctions | COL7A1 | Epidermolysis bullosa dystrophica |
| VIII | Some endothelial cells | COL8A1, COL8A2 | Posterior polymorphous corneal dystrophy 2 |
| IX | FACIT collagen, cartilage, assoc. with type II and XI fibrils | COL9A1, COL9A2, COL9A3 | EDM2 and EDM3 |
| X | Hypertrophic and mineralizing cartilage | COL10A1 | Schmid metaphyseal dysplasia |
| XI | Cartilage | COL11A1, COL11A2 | Collagenopathy, types II and XI |
| XII | FACIT collagen, interacts with type I containing fibrils, decorin and glycosaminoglycans | COL12A1 | – |
| XIII | Transmembrane collagen, interacts with integrin a1b1, fibronectin and components of basement membranes like nidogen and perlecan. | COL13A1 | – |
| XIV | FACIT collagen, also known as undulin | COL14A1 | – |
| XV | – | COL15A1 | – |
| XVI | – | COL16A1 | – |
| XVII | Transmembrane collagen, also known as BP180, a 180 kDa protein | COL17A1 | Bullous pemphigoid and certain forms of junctional epidermolysis bullosa |
| XVIII | Source of endostatin | COL18A1 | – |
| XIX | FACIT collagen | COL19A1 | – |
| XX | – | COL20A1 | – |
| XXI | FACIT collagen | COL21A1 | – |
| XXII | – | COL22A1 | – |
| XXIII | MACIT collagen | COL23A1 | – |
| XXIV | – | COL24A1 | – |
| XXV | – | COL25A1 | – |
| XXVI | – | EMID2 | – |
| XXVII | – | COL27A1 | – |
| XXVIII | – | COL28A1 | – |
| XXIX | Epidermal collagen | COL29A1 | Atopic dermatitis[41] |
In addition to the above-mentioned disorders, excessive deposition of collagen occurs in scleroderma.
Diseases
One thousand mutations have been identified in 12 out of more than 20 types of collagen. These mutations can lead to various diseases at the tissue level.[42]
Osteogenesis imperfecta – Caused by a mutation in type 1 collagen, dominant autosomal disorder, results in weak bones and irregular connective tissue, some cases can be mild while others can be lethal. Mild cases have lowered levels of collagen type 1 while severe cases have structural defects in collagen.[43]
Chondrodysplasias – Skeletal disorder believed to be caused by a mutation in type 2 collagen, further research is being conducted to confirm this.[44]
Ehlers-Danlos syndrome – Thirteen different types of this disorder, which lead to deformities in connective tissue, are known.[45] Some of the rarer types can be lethal, leading to the rupture of arteries. Each syndrome is caused by a different mutation. For example, the vascular type (vEDS) of this disorder is caused by a mutation in collagen type 3.[46]
Alport syndrome – Can be passed on genetically, usually as X-linked dominant, but also as both an autosomal dominant and autosomal recessive disorder, sufferers have problems with their kidneys and eyes, loss of hearing can also develop during the childhood or adolescent years.[47]
Knobloch syndrome – Caused by a mutation in the COL18A1 gene that codes for the production of collagen XVIII. Patients present with protrusion of the brain tissue and degeneration of the retina; an individual who has family members suffering from the disorder is at an increased risk of developing it themselves since there is a hereditary link.[42]
Characteristics
Collagen is one of the long, fibrous structural proteins whose functions are quite different from those of globular proteins, such as enzymes. Tough bundles of collagen called collagen fibers are a major component of the extracellular matrix that supports most tissues and gives cells structure from the outside, but collagen is also found inside certain cells. Collagen has great tensile strength, and is the main component of fascia, cartilage, ligaments, tendons, bone and skin.[48][49] Along with elastin and soft keratin, it is responsible for skin strength and elasticity, and its degradation leads to wrinkles that accompany aging.[11] It strengthens blood vessels and plays a role in tissue development. It is present in the cornea and lens of the eye in crystalline form. It may be one of the most abundant proteins in the fossil record, given that it appears to fossilize frequently, even in bones from the Mesozoic and Paleozoic.[50]
Uses

Collagen has a wide variety of applications, from food to medical. For instance, it is used in cosmetic surgery and burn surgery. It is widely used in the form of collagen casings for sausages.[51][52]
If collagen is subject to sufficient denaturation, e.g. by heating, the three tropocollagen strands separate partially or completely into globular domains, containing a different secondary structure to the normal collagen polyproline II (PPII), e.g. random coils. This process describes the formation of gelatin, which is used in many foods, including flavored gelatin desserts. Besides food, gelatin has been used in pharmaceutical, cosmetic, and photography industries. It is also used as a dietary supplement.
From the Greek for glue, kolla, the word collagen means "glue producer" and refers to the early process of boiling the skin and sinews of horses and other animals to obtain glue. Collagen adhesive was used by Egyptians about 4,000 years ago, and Native Americans used it in bows about 1,500 years ago. The oldest glue in the world, carbon-dated as more than 8,000 years old, was found to be collagen—used as a protective lining on rope baskets and embroidered fabrics, and to hold utensils together; also in crisscross decorations on human skulls.[53] Collagen normally converts to gelatin, but survived due to dry conditions. Animal glues are thermoplastic, softening again upon reheating, so they are still used in making musical instruments such as fine violins and guitars, which may have to be reopened for repairs—an application incompatible with tough, synthetic plastic adhesives, which are permanent. Animal sinews and skins, including leather, have been used to make useful articles for millennia.
Gelatin-resorcinol-formaldehyde glue (and with formaldehyde replaced by less-toxic pentanedial and ethanedial) has been used to repair experimental incisions in rabbit lungs.[54]
Adaptation and diversification
The evolution of collagens was a fundamental step in the early evolution of animals, supporting multicellular animal forms.[55] Collagens are the most abundant proteins in vertebrates, making up some 30% of all proteins in the human body.[56][57][58] Based on their molecular structures, collagen proteins are divided into two main classes – fibril-forming (or fibrillar) collagens and non-fibril-forming (non-fibrillar) collagens – which are further divided into 28 different types (as of 2017), based on individual structures and functions that the protein specifically has in the body.[57][58] Fibrillar collagen, producing the three-dimensional frameworks in different tissues and organs, derived from a single common ancestor during evolution.[57][59] Non-fibrillar collagen is the major supporting component of the extracellular matrix.[58]
The morphology of fibrillar and non-fibrillar collagen types became differentiated during divergent evolution.[58] These two collagen types arose from different mutation and gene duplication events that evolved to the current 28 types of collagen proteins providing the diversification of collagen-supporting structures in the body, such as the skeleton, which formed from the alpha (denoted as α) collagen gene.[57][59] Fibrillar collagen was co-opted during evolutionary adaptation from existing genes by natural selection to construct new organ and tissue structures, enabling the emergence of evolved species with improved capabilities.[57][59]
History
The molecular and packing structures of collagen have eluded scientists over decades of research. The first evidence that it possesses a regular structure at the molecular level was presented in the mid-1930s.[60][61] Since that time, research concentrated on the conformation of the collagen monomer, producing several competing models, although correctly dealing with the conformation of each individual peptide chain. The triple-helical "Madras" model provided an accurate model of quaternary structure in collagen.[62][63][64] This model was supported by further studies showing higher resolution in the late 20th century.[65][66][67][68]
The packing structure of collagen has not been defined to the same degree outside of the fibrillar collagen types, although it has been long known to be hexagonal.[33][69][70] As with its monomeric structure, several conflicting models alleged that either the packing arrangement of collagen molecules is 'sheet-like' or microfibrillar.[71][72] The microfibrillar structure of collagen fibrils in tendon, cornea and cartilage was imaged directly by electron microscopy in the late 20th century and early 21st century.[73][74][75] The microfibrillar structure of tail tendon was modeled as being closest to the observed structure, although it oversimplified the topological progression of neighboring collagen molecules, and so did not predict the correct conformation of the discontinuous D-periodic pentameric arrangement termed microfibril.[32][76][77]
See also
- Animal glue
- Braising
- Collagen hybridizing peptide, a peptide that can bind to denatured collagen
- Collagenase, the enzyme involved in collagen breakdown and remodeling
- Fibrous protein
- Gelatin
- Hydrolyzed collagen, a common form in which collagen is sold as a supplement
- Hypermobility syndrome
- Lysyl oxidase
- MMP inhibitors
- Osteoid, collagen-containing component of bone
References
- Di Lullo, Gloria A.; Sweeney, Shawn M.; Körkkö, Jarmo; Ala-Kokko, Leena & San Antonio, James D. (2002). "Mapping the Ligand-binding Sites and Disease-associated Mutations on the Most Abundant Protein in the Human, Type I Collagen". J. Biol. Chem. 277 (6): 4223–4231. doi:10.1074/jbc.M110709200. PMID 11704682.
- "Leather grown using biotechnology is about to hit the catwalk". The Economist. 26 August 2017. Archived from the original on 1 September 2017. Retrieved 2 September 2017.
- Britannica Concise Encyclopedia 2007
- Sikorski, Zdzisław E. (2001). Chemical and Functional Properties of Food Proteins. Boca Raton: CRC Press. p. 242. ISBN 978-1-56676-960-0.
- Bogue, Robert H. (1923). "Conditions Affecting the Hydrolysis of Collagen to Gelatin". Industrial and Engineering Chemistry. 15 (11): 1154–1159. doi:10.1021/ie50167a018.
- O.E.D. 2nd Edition 2005
- Müller, Werner E. G. (2003). "The Origin of Metazoan Complexity: Porifera as Integrated Animals". Integrative and Comparative Biology. 43 (1): 3–10. CiteSeerX 10.1.1.333.3174. doi:10.1093/icb/43.1.3. PMID 21680404. S2CID 17232196.
- Sabiston textbook of surgery board review, 7th edition. Chapter 5 wound healing, question 14
- Ricard-Blum, S. (2011). "The Collagen Family". Cold Spring Harbor Perspectives in Biology. 3 (1): a004978. doi:10.1101/cshperspect.a004978. PMC 3003457. PMID 21421911.
- Franzke, CW; Bruckner, P; Bruckner-Tuderman, L (11 February 2005). "Collagenous transmembrane proteins: recent insights into biology and pathology". The Journal of Biological Chemistry. 280 (6): 4005–8. doi:10.1074/jbc.R400034200. PMID 15561712.
- Dermal Fillers | The Ageing Skin Archived 13 May 2011 at the Wayback Machine. Pharmaxchange.info. Retrieved on 21 April 2013.
- Cunniffe, G; F O'Brien (2011). "Collagen scaffolds for orthopedic regenerative medicine". The Journal of the Minerals, Metals and Materials Society. 63 (4): 66–73. Bibcode:2011JOM....63d..66C. doi:10.1007/s11837-011-0061-y. S2CID 136755815.
- Geiger, M (2003). "Collagen sponges for bone regeneration with rhBMP-2". Advanced Drug Delivery Reviews. 55 (12): 1613–1629. doi:10.1016/j.addr.2003.08.010. ISSN 0169-409X.
- Bunyaratavej, Pintippa; Wang, Hom-Lay (2001). "Collagen Membranes: A Review". Journal of Periodontology. 72 (2): 215–229. doi:10.1902/jop.2001.72.2.215. hdl:2027.42/141506. ISSN 0022-3492.
- Drury, Jeanie L.; Mooney, David J. (2003). "Hydrogels for tissue engineering: scaffold design variables and applications". Biomaterials. 24 (24): 4337–4351. doi:10.1016/S0142-9612(03)00340-5. ISSN 0142-9612.
- Tonndorf, Robert; Aibibu, Dilbar; Cherif, Chokri (2020). "Collagen multifilament spinning". Materials Science and Engineering: C. 106: 110105. doi:10.1016/j.msec.2019.110105. ISSN 0928-4931.
- Oliveira, S; R Ringshia; R Legeros; E Clark; L Terracio; C Teixeira M Yost (2009). "An improved collagen scaffold for skeletal regeneration". Journal of Biomedical Materials. 94 (2): 371–379. doi:10.1002/jbm.a.32694. PMC 2891373. PMID 20186736.
- Onkar, Singh; Gupta, Shilpi Singh; Soni, Mohan; Moses, Sonia; Shukla, Sumit; Mathur, Raj Kumar (January 2011). "Collagen Dressing Versus Conventional Dressings in Burn and Chronic Wounds: A Retrospective Study". Journal of Cutaneous and Aesthetic Surgery. 4 (1): 12–16. doi:10.4103/0974-2077.79180. PMC 3081477. PMID 21572675.
- Gould, L. J. (2016). "Topical Collagen-Based Biomaterials for Chronic Wounds: Rationale and Clinical Application". Advances in Wound Care. 5 (1): 19–31. doi:10.1089/wound.2014.0595. PMC 4717516. PMID 26858912.
- Birbrair, Alexander; Zhang, Tan; Files, Daniel C.; Mannava, Sandeep; Smith, Thomas; Wang, Zhong-Min; Messi, Maria L.; Mintz, Akiva; Delbono, Osvaldo (6 November 2014). "Type-1 pericytes accumulate after tissue injury and produce collagen in an organ-dependent manner". Stem Cell Research & Therapy. 5 (6): 122. doi:10.1186/scrt512. ISSN 1757-6512. PMC 4445991. PMID 25376879.
- Singh, O; SS Gupta; M Soni; S Moses; S Shukla; RK Mathur (2011). "Collagen dressing versus conventional dressings in burn and chronic wounds: a retrospective study". Journal of Cutaneous and Aesthetic Surgery. 4 (1): 12–16. doi:10.4103/0974-2077.79180. PMC 3081477. PMID 21572675.
- Blow, Nathan (2009). "Cell culture: building a better matrix". Nature Methods. 6 (8): 619–622. doi:10.1038/nmeth0809-619. S2CID 33438539.
- Szpak, Paul (2011). "Fish bone chemistry and ultrastructure: implications for taphonomy and stable isotope analysis". Journal of Archaeological Science. 38 (12): 3358–3372. doi:10.1016/j.jas.2011.07.022.
- Peterkofsky, B (1991). "Ascorbate requirement for hydroxylation and secretion of procollagen: Relationship to inhibition of collagen synthesis in scurvy". American Journal of Clinical Nutrition. 54 (6 Suppl): 1135S–1140S. doi:10.1093/ajcn/54.6.1135s. PMID 1720597.
- Gorres, K. L.; Raines, R. T. (2010). "Prolyl 4-hydroxylase". Crit. Rev. Biochem. Mol. Biol. 45 (2): 106–24. doi:10.3109/10409231003627991. PMC 2841224. PMID 20199358.
- Myllylä, R.; Majamaa, K.; Günzler, V.; Hanauske-Abel, H. M.; Kivirikko, K. I. (1984). "Ascorbate is consumed stoichiometrically in the uncoupled reactions catalyzed by propyl 4-hydroxylase and lysyl hydroxylase". J. Biol. Chem. 259 (9): 5403–5. PMID 6325436.
- Houck, J. C.; Sharma, V. K.; Patel, Y. M.; Gladner, J. A. (1968). "Induction of Collagenolytic and Proteolytic Activities by AntiInflammatory Drugs in the Skin and Fibroblasts". Biochemical Pharmacology. 17 (10): 2081–2090. doi:10.1016/0006-2952(68)90182-2. PMID 4301453.
- "preprocollagen". The Free Dictionary.
- Al-Hadithy, H.; Isenberg, DA; et al. (1982). "Neutrophil function in systemic lupus erythematosus and other collagen diseases". Ann Rheum Dis. 41 (1): 33–38. doi:10.1136/ard.41.1.33. PMC 1000860. PMID 7065727.
- Hulmes, D. J. (2002). "Building collagen molecules, fibrils, and suprafibrillar structures". J Struct Biol. 137 (1–2): 2–10. doi:10.1006/jsbi.2002.4450. PMID 12064927.
- Hulmes, D. J. (1992). "The collagen superfamily—diverse structures and assemblies". Essays Biochem. 27: 49–67. PMID 1425603.
- Orgel, J. P.; Irving, TC; et al. (2006). "Microfibrillar structure of type I collagen in situ". PNAS. 103 (24): 9001–9005. Bibcode:2006PNAS..103.9001O. doi:10.1073/pnas.0502718103. PMC 1473175. PMID 16751282.
- Hulmes, D. J. & Miller, A. (1979). "Quasi-hexagonal molecular packing in collagen fibrils". Nature. 282 (5741): 878–880. Bibcode:1979Natur.282..878H. doi:10.1038/282878a0. PMID 514368. S2CID 4332269.
- Perumal, S.; Antipova, O. & Orgel, J. P. (2008). "Collagen fibril architecture, domain organization, and triple-helical conformation govern its proteolysis". PNAS. 105 (8): 2824–2829. Bibcode:2008PNAS..105.2824P. doi:10.1073/pnas.0710588105. PMC 2268544. PMID 18287018.
- Buchanan, Jenna K.; Zhang, Yi; Holmes, Geoff; Covington, Anthony D.; Prabakar, Sujay (2019). "Role of X-ray Scattering Techniques in Understanding the Collagen Structure of Leather". ChemistrySelect. 4 (48): 14091–14102. doi:10.1002/slct.201902908. ISSN 2365-6549.
- Sweeney, S. M.; Orgel, JP; et al. (2008). "Candidate Cell and Matrix Interaction Domains on the Collagen Fibril, the Predominant Protein of Vertebrates". J Biol Chem. 283 (30): 21187–21197. doi:10.1074/jbc.M709319200. PMC 2475701. PMID 18487200.
- Twardowski, T.; Fertala, A.; et al. (2007). "Type I collagen and collagen mimetics as angiogenesis promoting superpolymers". Curr Pharm Des. 13 (35): 3608–3621. doi:10.2174/138161207782794176. PMID 18220798.
- Minary-Jolandan, M; Yu, MF (2009). "Nanomechanical heterogeneity in the gap and overlap regions of type I collagen fibrils with implications for bone heterogeneity". Biomacromolecules. 10 (9): 2565–70. doi:10.1021/bm900519v. PMID 19694448.
- Ross, M. H. and Pawlina, W. (2011) Histology, 6th ed., Lippincott Williams & Wilkins, p. 218.
- Söderhäll, C.; Marenholz, I.; Kerscher, T.; Rüschendorf, F; Rüschendorf, F.; Esparza-Gordillo, J.; Mayr, G; et al. (2007). "Variants in a Novel Epidermal Collagen Gene (COL29A1) Are Associated with Atopic Dermatitis". PLOS Biology. 5 (9): e242. doi:10.1371/journal.pbio.0050242. PMC 1971127. PMID 17850181.
- "Collagen Types and Linked Disorders". News-Medical.net. 18 January 2011. Archived from the original on 1 December 2017. Retrieved 19 November 2017.
- Mahajan VB, Olney AH, Garrett P, Chary A, Dragan E, Lerner G, Murray J, Bassuk AG (2010). "Collagen XVIII mutation in Knobloch syndrome with acute lymphoblastic leukemia". American Journal of Medical Genetics Part A. 152A (11): 2875–9. doi:10.1002/ajmg.a.33621. PMC 2965270. PMID 20799329.
- Gajko-Galicka, A (2002). "Mutations in type I collagen genes resulting in osteogenesis imperfecta in humans" (PDF). Acta Biochimica Polonica. 49 (2): 433–41. doi:10.18388/abp.2002_3802. PMID 12362985. Archived (PDF) from the original on 7 June 2013.
- Horton WA, Campbell D, Machado MA, Chou J (1989). "Type II collagen screening in the human chondrodysplasias". Am. J. Med. Genet. 34 (4): 579–83. doi:10.1002/ajmg.1320340425. PMID 2624272.
- Malfait F, Francomano C, Byers P, Belmont J, Berglund B, Black J, Bloom L, Bowen JM, Brady AF, Burrows NP, Castori M, Cohen H, Colombi M, Demirdas S, De Backer J, De Paepe A, Fournel-Gigleux S, Frank M, Ghali N, Giunta C, Grahame R, Hakim A, Jeunemaitre X, Johnson D, Juul-Kristensen B, Kapferer-Seebacher I, Kazkaz H, Kosho T, Lavallee ME, Levy H, Mendoza-Londono R, Pepin M, Pope FM, Reinstein E, Robert L, Rohrbach M, Sanders L, Sobey GJ, Van Damme T, Vandersteen A, van Mourik C, Voermans N, Wheeldon N, Zschocke J, Tinkle B. 2017. The 2017 international classification of the Ehlers–Danlos syndromes. Am J Med Genet Part C Semin Med Genet 175C:8–26.
- Hamel BC, Pals G, Engels CH, van den Akker E, Boers GH, van Dongen PW, Steijlen PM (1998). "Ehlers-Danlos syndrome and type III collagen abnormalities: a variable clinical spectrum". Clin. Genet. 53 (6): 440–6. doi:10.1111/j.1399-0004.1998.tb02592.x. PMID 9712532. S2CID 39089732.
- Kashtan, CE (1993) "Collagen IV-Related Nephropathies (Alport Syndrome and Thin Basement Membrane Nephropathy Archived 2017-12-25 at the Wayback Machine)", in RA Pagon, TD Bird, CR Dolan, K Stephens & MP Adam (eds), GeneReviews, University of Washington, Seattle, Seattle WA PMID 20301386.
- Fratzl, P. (2008). Collagen: Structure and Mechanics. New York: Springer. ISBN 978-0-387-73905-2.
- Buehler, M. J. (2006). "Nature designs tough collagen: Explaining the nanostructure of collagen fibrils". PNAS. 103 (33): 12285–12290. Bibcode:2006PNAS..10312285B. doi:10.1073/pnas.0603216103. PMC 1567872. PMID 16895989.
- Zylberberg, L.; Laurin, M. (2011). "Analysis of fossil bone organic matrix by transmission electron microscopy". Comptes Rendus Palevol. 11 (5–6): 357–366. doi:10.1016/j.crpv.2011.04.004.
- https://www.lemproducts.com/understanding_collagen_casings
- http://askthemeatman.com/what_are_collagen_casings.htm
- Walker, Amélie A. (21 May 1998). "Oldest Glue Discovered". Archaeology. Archived from the original on 17 December 2005.
- Ennker, I. C.; Ennker, JüRgen; et al. (1994). "Formaldehyde-free collagen glue in experimental lung gluing". Ann. Thorac. Surg. 57 (6): 1622–1627. doi:10.1016/0003-4975(94)90136-8. PMID 8010812. Archived from the original on 8 July 2012.
- Exposito, J. Y.; Cluzel, C; Garrone, R; Lethias, C (2002). "Evolution of collagens". The Anatomical Record. 268 (3): 302–16. doi:10.1002/ar.10162. PMID 12382326. S2CID 12376172.
- Karsdal, Morten (2016). Biochemistry of collagens: structure, function and biomarkers. London, United Kingdom: Elsevier Science. ISBN 978-0-12-809899-8. OCLC 955139533.
- Ricard-Blum, Sylvie (2011). "The collagen family (Review)". Cold Spring Harbor Perspectives in Biology. 3 (1). doi:10.1101/cshperspect.a004978. ISSN 1943-0264. PMC 3003457. PMID 21421911.
- Bella, Jordi; Hulmes, David J. S. (2017). "Fibrillar collagens". Subcellular Biochemistry. 82: 457–490. doi:10.1007/978-3-319-49674-0_14. ISBN 978-3-319-49672-6. ISSN 0306-0225. PMID 28101870.
- Rodriguez-Pascual, Fernando; Slatter, David Anthony (23 November 2016). "Collagen cross-linking: insights on the evolution of metazoan extracellular matrix". Scientific Reports. 6 (1): 37374. Bibcode:2016NatSR...637374R. doi:10.1038/srep37374. ISSN 2045-2322. PMC 5120351. PMID 27876853.
- Wyckoff, R.; Corey, R. & Biscoe, J. (1935). "X-ray reflections of long spacing from tendon". Science. 82 (2121): 175–176. Bibcode:1935Sci....82..175W. doi:10.1126/science.82.2121.175. PMID 17810172.
- Clark, G.; Parker, E.; Schaad, J. & Warren, W. J. (1935). "New measurements of previously unknown large interplanar spacings in natural materials". J. Am. Chem. Soc. 57 (8): 1509. doi:10.1021/ja01311a504.
- Balasubramanian, D . (October 2001). "GNR — A Tribute". Resonance. 6 (10): 2–4. doi:10.1007/BF02836961. S2CID 122261106. Archived from the original on 10 January 2014.
- Leonidas, Demetres D.; Chavali, GB; et al. (2001). "Binding of phosphate and pyrophosphate ions at the active site of human angiogenin as revealed by X-ray crystallography". Protein Science. 10 (8): 1669–1676. doi:10.1110/ps.13601. PMC 2374093. PMID 11468363.
- Subramanian, Easwara (2001). "Obituary: G.N. Ramachandran". Nature Structural & Molecular Biology. 8 (6): 489–491. doi:10.1038/88544. PMID 11373614. S2CID 7231304.
- Fraser, R. D.; MacRae, T. P. & Suzuki, E. (1979). "Chain conformation in the collagen molecule". J Mol Biol. 129 (3): 463–481. doi:10.1016/0022-2836(79)90507-2. PMID 458854.
- Okuyama, K.; Okuyama, K; et al. (1981). "Crystal and molecular structure of a collagen-like polypeptide (Pro-Pro-Gly)10". J Mol Biol. 152 (2): 427–443. doi:10.1016/0022-2836(81)90252-7. PMID 7328660.
- Traub, W.; Yonath, A. & Segal, D. M. (1969). "On the molecular structure of collagen". Nature. 221 (5184): 914–917. Bibcode:1969Natur.221..914T. doi:10.1038/221914a0. PMID 5765503. S2CID 4145093.
- Bella, J.; Eaton, M.; Brodsky, B.; Berman, H. M. (1994). "Crystal and molecular structure of a collagen-like peptide at 1.9 A resolution". Science. 266 (5182): 75–81. Bibcode:1994Sci...266...75B. doi:10.1126/science.7695699. PMID 7695699.
- Jesior, J. C.; Miller, A. & Berthet-Colominas, C. (1980). "Crystalline three-dimensional packing is general characteristic of type I collagen fibrils". FEBS Lett. 113 (2): 238–240. doi:10.1016/0014-5793(80)80600-4. PMID 7389896. S2CID 40958154.
- Fraser, R. D. B. & MacRae, T. P. (1981). "Unit cell and molecular connectivity in tendon collagen". Int. J. Biol. Macromol. 3 (3): 193–200. doi:10.1016/0141-8130(81)90063-5.
- Fraser, R. D.; MacRae, T. P.; Miller, A. (1987). "Molecular packing in type I collagen fibrils". J Mol Biol. 193 (1): 115–125. doi:10.1016/0022-2836(87)90631-0. PMID 3586015.
- Wess, T. J.; Hammersley, AP; et al. (1998). "Molecular packing of type I collagen in tendon". J Mol Biol. 275 (2): 255–267. doi:10.1006/jmbi.1997.1449. PMID 9466908.
- Raspanti, M.; Ottani, V.; Ruggeri, A. (1990). "Subfibrillar architecture and functional properties of collagen: a comparative study in rat tendons". J. Anat. 172: 157–164. PMC 1257211. PMID 2272900.
- Holmes, D. F.; Gilpin, C. J.; Baldock, C.; Ziese, U.; Koster, A. J.; Kadler, K. E. (2001). "Corneal collagen fibril structure in three dimensions: Structural insights into fibril assembly, mechanical properties, and tissue organization". PNAS. 98 (13): 7307–7312. Bibcode:2001PNAS...98.7307H. doi:10.1073/pnas.111150598. PMC 34664. PMID 11390960.
- Holmes, D. F.; Kadler, KE (2006). "The 10+4 microfibril structure of thin cartilage fibrils". PNAS. 103 (46): 17249–17254. Bibcode:2006PNAS..10317249H. doi:10.1073/pnas.0608417103. PMC 1859918. PMID 17088555.
- Okuyama, K; Bächinger, HP; Mizuno, K; Boudko, SP; Engel, J; Berisio, R; Vitagliano, L (2009). "Comment on Microfibrillar structure of type I collagen in situ by Orgel et al. (2006), Proc. Natl Acad. Sci. USA, 103, 9001–9005". Acta Crystallographica Section D. 65 (Pt9): 1009–10. doi:10.1107/S0907444909023051. PMID 19690380.
- Orgel, Joseph (2009). "On the packing structure of collagen: response to Okuyama et al.'s comment on Microfibrillar structure of type I collagen in situ". Acta Crystallographica Section D. D65 (9): 1009. doi:10.1107/S0907444909028741.
| Wikimedia Commons has media related to Collagen. |