Haplogroup E-M2
Haplogroup E-M2 is a human Y-chromosome DNA haplogroup. It is primarily distributed in Sub-Saharan Africa. E-M2 is the predominant subclade in Western Africa, Central Africa, Southern Africa and the African Great Lakes, and occurs at moderate frequencies in North Africa and Middle East. E-M2 has several subclades, but many of these subhaplogroups are included in either E-L485 or E-U175. E-M2 is especially common in native Africans speaking Niger-Congo languages and was spread to Southern and Eastern Africa through the Bantu expansion.
| Haplogroup E-M2 (former E3a / E1b1a) | |
|---|---|
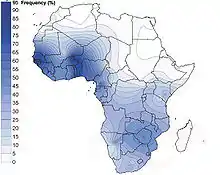
.png.webp) Interpolated frequency distribution.[1] | |
| Possible time of origin | 39,300 years BP[2] |
| Coalescence age | 15,700 years BP[2] |
| Possible place of origin | Western Africa[3][4] or Central Africa[3][4] |
| Ancestor | E-V38 |
| Descendants | E-Z5994, E-V43 |
| Defining mutations | M2, DYS271/SY81, M291, P1/PN1, P189.1, P293.1 |
Origins
The discovery of two SNPs (V38 and V100) by Trombetta et al. (2011) significantly redefined the E-V38 phylogenetic tree. This led the authors to suggest that E-V38 may have originated in East Africa. E-V38 joins the West African-affiliated E-M2 and the northern East African-affiliated E-M329 with an earlier common ancestor who, like E-P2, may have also originated in East Africa.[5] The downstreams SNP E-M180 possibly originated on the moist south-central Saharan savannah/grassland of northern Africa between 14,000–10,000 years BP.[6][7][8][9] According to Wood et al. (2005) and Rosa et al. (2007), such population movements changed the pre-existing population Y chromosomal diversity in Central, Southern and southern East Africa, replacing the previous haplogroups frequencies in these areas with the now dominant E1b1a1 lineages. Traces of earlier inhabitants, however, can be observed today in these regions via the presence of the Y DNA haplogroups A1a, A1b, A2, A3, and B-M60 that are common in certain populations, such as the Mbuti and Khoisan.[10][11][12]
Distribution
This haplogroup's frequency and diversity are highest in the West Africa region. Within Africa, E-M2 displays a west-to-east as well as a south-to-north clinal distribution. In other words, the frequency of the haplogroup decreases as one moves from western and southern Africa toward the eastern and northern parts of the continent.[13]
| Population group | frequency | References |
|---|---|---|
| Bamileke | 96%-100% | [13][14] |
| Ewe | 97% | [11] |
| Ga | 97% | [11] |
| Yoruba | 93.1% | [15] |
| Tutsi | 85% | [13] |
| Fante | 84% | [11] |
| Mandinka | 79%–87% | [10][11] |
| Ovambo | 82% | [11] |
| Senegalese | 81% | [16] |
| Ganda | 77% | [11] |
| Bijagós | 76% | [10] |
| Balanta | 73% | [10] |
| Fula | 73% | [10] |
| Herero | 71% | [11] |
| Nalú | 71% | [10] |
Populations on the North West Africa, central Eastern Africa and Madagascar have tested at more moderate frequencies.
| Population group | frequency | References |
|---|---|---|
| Tuareg from Tânout, Niger | 44.4% (8/18 subjects) | [17] |
| Comorian Shirazi | 41% | [18] |
| Tuareg from Gorom-Gorom, Burkina Faso | 16.6% (3/18) | [17] |
| Tuareg from Gossi, Mali | 9.1% (1/9) | [17] |
| Cape Verdeans | 15.9% (32/201) | [19] |
| Maasai | 15.4% (4/26) | [11] |
| Luo | 66% (6/9) | [11] |
| Iraqw | 11.11% (1/9) | [11] |
| Comoros | 23.46% (69/294) | [18] |
| Merina people (also called Highlanders) | 44% (4/9) | [20] |
| Antandroy | 69.6% (32/46) | [20] |
| Antanosy | 48.9% (23/47) | [20] |
| Antaisaka | 37.5% (3/8) | [20] |
E-M2 is found at low to moderate frequencies in North Africa, and northern East Africa. Some of the lineages found in these areas are possibly due to the Bantu expansion or other migrations.[13][21] The E-M2 marker that appeared in North African samples stem from Haratin (an ethnic group of black African origin indigenous to northwest Africa, related to but distinct from Sub-Saharan West Africans).[13] However, the discovery in 2011 of the E-M2 marker that predates E-M2 has led Trombetta et al. to suggest that E-M2 may have originated in East Africa (please refer to the Origins section for details). In Eritrea and most of Ethiopia (excluding the Anuak) E-V38 is usually only found in the form of E-M329, which is autochthonous, while E-M2 generally indicates Bantu migratory origins.[22][23][24]
| Population group | frequency | References |
|---|---|---|
| Tuareg from Al Awaynat and Tahala, Libya | 46.5% (20/43) [Note 1] | [25] |
| Oran, Algeria | 8.6% (8/93) | [26] |
| Berbers, southern and north-central Morocco | 9.5% (6/63) 5.8% (4/69) | [27][Note 2][28] |
| Moroccan Arabs | 6.8% (3/44) 1.9% (1/54) | [27][28] |
| Saharawis | 3.5% (1/29) | [27] |
| Egyptians | 1.4% (2/147), 0% (0/73), 8.33% (3/36) | [13][29][30] |
| Tunisians | 1.4% (2/148) | [30] |
| Sudanese (may include Hausa migrants) | 0.9% (4/445) | [31] |
| Somalia nationals (may include Bantu minorities) | 1.5% (3/201) | [21] |
| Somalis (ethnic Somalis) | 0% (0/108) | [24] |
| Djiboutians (Somalis and Afars) | 0% (0/54) | [22] |
| Eritreans (Tigray-Tigrinya, Tigre, Cunama, Nara, Saho) | 0% (0/161)[Note 3] | [22] |
| Ethiopians (Amhara, Oromo, Ethiopian Jews, Wolayta, Somali, Tigray, Other Ethiopians) | 0% (0/119)[Note 4] | [22] |
Outside of Africa, E-M2 has been found at low frequencies. The clade has been found at low frequencies in West Asia. A few isolated occurrences of E-M2 have also been observed among populations in Southern Europe, such as Croatia, Malta, Spain and Portugal.[32] [33][34][35]
| Population group | frequency | References |
|---|---|---|
| Saudi Arabians | 6.6% (11/157) | |
| Omanis | 6.6% (8/121) | [13] |
| Emiratis | 5.5% (9/164) | [37] |
| Yemenis | 4.8% (3/62) | [37] |
| Cypriots | 3.2% (2/62) | [35] |
| Southern Iranians | 1.7% (2/117) | [38] |
| Jordanians | 1.4% (2/139) | [39] |
| Sri Lanka | 1.4% (9/638) | [40] |
| Aeolian Islands, Italy | 1.2% (1/81) | [41] |
The Trans-Atlantic slave trade brought people to North America, Central America and South America including the Caribbean. Consequently, the haplogroup is often observed in the United States populations in men who self-identify as African Americans.[42] It has also been observed in a number of populations in Mexico, the Caribbean, Central America, and South America among people of African descent.
| Population group | frequency | References |
|---|---|---|
| Americans | 7.7–7.9% [Note 5] | [42] |
| Cubans | 9.8% (13/132) | [43] |
| Dominicans | 5.69% (2/26) | [44] |
| Puerto Ricans | 19.23% (5/26) | [44] |
| Nicaraguans | 5.5% (9/165) | [45] |
| Several populations of Colombians | 6.18% (69/1116) | [46] |
| Alagoas, Brazil | 4.45% (11/247) | [47] |
| Bahia, Brazil | 19% (19/100) | [48] |
| Bahamians | 58.63% (251/428) | [49] |
Subclades
E1b1a1
E1b1a1 is defined by markers DYS271/M2/SY81, M291, P1/PN1, P189, P293, V43, and V95. Whilst E1b1a reaches its highest frequency of 81% in Senegal, only 1 of the 139 Senegalese that were tested showed M191/P86.[15] In other words, as one moves to West Africa from western Central Africa, the less subclade E1b1a1f is found. "A possible explanation might be that haplotype 24 chromosomes [E-M2*] were already present across the Sudanese belt when the M191 mutation, which defines haplotype 22, arose in central western Africa. Only then would a later demic expansion have brought haplotype 22 chromosomes from central western to western Africa, giving rise to the opposite clinal distributions of haplotypes 22 and 24."[13]
E1b1a1a1
E1b1a1a1 is commonly defined by M180/P88. The basal subclade is quite regularly observed in M2+ samples.
E1b1a1a1a
E1b1a1a1a is defined by marker M58. 5% (2/37) of the town Singa-Rimaïbé, Burkina Faso tested positive for E-M58.[14] 15% (10/69) of Hutus in Rwanda tested positive for M58.[13] Three South Africans tested positive for this marker.[12] One Carioca from Rio de Janeiro, Brazil tested positive for the M58 SNP.[50] The place of origin and age is unreported.
E1b1a1a1b
E1b1a1a1b is defined by M116.2, a private marker. A single carrier was found in Mali.[12] [Note 6]
E1b1a1a1c
E1b1a1a1c is defined by private marker M149. This marker was found in a single South African.[12]
E1b1a1a1d
E1b1a1a1d is defined by a private marker M155. It is known from a single carrier in Mali.[12]
E1b1a1a1e
E1b1a1a1e is defined by markers M10, M66, M156 and M195. Wairak people in Tanzania tested 4.6% (2/43) positive for E-M10.[13] E-M10 was found in a single person of the Lissongo group in the Central African Republic and two members in a "Mixed" population from the Adamawa region.[12]
E1b1a1a1f
E1b1a1a1f is defined by L485. The basal node E-L485* appears to be somewhat uncommon but has not been sufficiently tested in large populations. The ancestral L485 SNP (along with several of its subclades) was very recently discovered. Some of these SNPs have little or no published population data and/or have yet to receive nomenclature recognition by the YCC.
- E1b1a1a1f1 is defined by marker L514. This SNP is currently without population study data outside of the 1000 Genomes Project.
- E1b1a1a1f1a (YCC E1b1a7) is defined by marker M191/P86. Filippo et al. (2011) studied a number of African populations that were E-M2 positive and found the basal E-M191/P86 (without E-P252/U174) in a population of Gur speakers in Burkina Faso.[51] Montano et al. (2011) found similar sparse distribution of E-M191* in Nigeria, Gabon, Cameroon and Congo.[9] M191/P86 positive samples occurred in tested populations of Annang (38.3%), Ibibio (45.6%), Efik (45%), and Igbo (54.3%) living in Nigeria, West Africa.[52] E-M191/P86 appears in varying frequencies in Central and Southern Africa but almost all are also positive for P252/U174. Bantu-speaking South Africans (89/343) tested 25.9% positive and Khoe-San speaking South Africans tested 7.7% (14/183) positive for this SNP.[53] It also appears commonly in Africans living in the Americas. A population in Rio de Janeiro, Brazil tested 9.2% (12/130) positive.[50] 34.9% (29/83) of American Haplogroup E men tested positive for M191.[42]
- Veeramah et al. (2010) studies of the recombining portions of M191 positive Y chromosomes suggest that this lineage has "diffusely spread with multiple high frequency haplotypes implying a longer evolutionary period since this haplogroup arose".[52] The subclade E1b1a1a1f1a appears to express opposite clinal distributions to E1b1a1* in the West African Savanna region. Haplogroup E1b1a1a1f1a (E-M191) has a frequency of 23% in Cameroon (where it represents 42% of haplotypes carrying the DYS271 mutation or E-M2), 13% in Burkina Faso (16% of haplotypes carrying the M2/DYS271 mutation) and only 1% in Senegal.[16] Similarly, while E1b1a reaches its highest frequency of 81% in Senegal, only 1 of the 139 Senegalese that were tested showed M191/P86.[16] In other words, as one moves to West Africa from western Central Africa, the less subclade E1b1a1f is found. "A possible explanation might be that haplotype 24 chromosomes [E-M2*] were already present across the Sudanese belt when the M191 mutation, which defines haplotype 22, arose in central western Africa. Only then would a later demic expansion have brought haplotype 22 chromosomes from central western to western Africa, giving rise to the opposite clinal distributions of haplotypes 22 and 24."[14]
- E1b1a1a1f1a1 (YCC E1b1a7a) is defined by P252/U174. It appears to be the most common subclade of E-L485. It is believed to have originated near western Central Africa. It is rarely found in the most western portions of West Africa. Montano et al. (2011) found this subclade very prevalent in Nigeria and Gabon.[9] Filippo et al. (2011) estimated a tMRCA of ~4.2 kya from sample of Yoruba population positive for the SNP.[51]
- E1b1a1a1f1a1b (YCC E1b1a7a2) is defined by P115. This subclade has only been observed amongst Fang people of Central Africa.[9]
- E1b1a1a1f1a1c (YCC E1b1a7a3) is defined by P116. Montano et al. (2011) observed this SNP only in Gabon and a Bassa population from Cameroon.[9]
- E1b1a1a1f1a1d is defined by Z1704. This subclade has been observed across Africa. The 1000 Genomes Project Consortium found this SNP in Yoruba Nigerian, three Kenyan Luhyas and one African descent Puerto Rican.[54]
E1b1a1a1g
E1b1a1a1g (YCC E1b1a8) is defined by marker U175. The basal E-U175* is extremely rare. Montano et al. (2011) only found one out of 505 tested African subjects who was U175 positive but negative for U209.[9] Brucato et al. found similarly low frequencies of basal E-U175* in subjects in the Ivory Coast and Benin. Veeramah et al. (2010) found U175 in tested Annang (45.3%), Ibibio (37%), Efik (33.3%), and Igbo (25.3%) but did not test for U209.[52]
The supposed "Bantu haplotype" found in E-U175 carriers is "present at appreciable frequencies in other Niger–Congo languages speaking peoples as far west as Guinea-Bissau".[52] This is the modal haplotype of STR markers that is common in carriers of E-U175.[Note 7]
| E-U175 haplotype | DYS19 | DYS388 | DYS390 | DYS391 | DYS392 | DYS393 | |
| 15 | 12 | 21 | 10 | 11 | 13 | ||
E1b1a1a1g has several subclades.
- E1b1a1a1g1 (YCC E1b1a8a) is defined by U209. It is the most prominent subclade of U175. This subclade has very high frequencies of over fifty percentages in Cameroonian populations of Bassa and Bakaka, possibly indicating place of origin. However, E-U209 is widely found at lower frequencies in West and Central African countries surrounding Cameroon and Gabon.[9] Brucato et al. (2010) found the SNP in a populations of Ahizi (in Ivory Coast) 38.8% (19/49), Yacouba (Ivory Coast) 27.5% (11/40), and Beninese 6.5% (5/77) respectively.[56]
- E1b1a1a1g1a (YCC E1b1a8a1) is defined by U290. The Montano et al. (2011) study of U290 showed a lower frequency in Nigeria (11.7%) and western Central Africa than basal node U209. The highest population frequency rate in that study was 57.7% (15/26) in Ewondo in Cameroon.[9] 32.5% (27/83) of American Haplogroup E men tested by Sims et al. (2007) were positive for this SNP.[42]
- E1b1a1a1g1a2 is defined by Z1725. This marker has been observed by The 1000 Genomes Project Consortium in Yoruba Nigerians and Luhya Kenyans.[54]
- E1b1a1a1g1c (YCC E1b1a4) is defined by M154. A Bamilike population tested 31.3% (15/48) for the marker. Bakaka speakers from Cameroon tested 8%.[14] An Ovimbundu test population found this SNP at 14% (14/100).[57] Members of this subclade have also been found in South Africa.[58][53]
- E1b1a1a1g1d is defined by V39. Trombetta et al. first published this SNP in 2011 but gave little population data about it.[5] It is only known to have been found in an African population.
Phylogenetics
Phylogenetic history
Prior to 2002, there were in academic literature at least seven naming systems for the Y-Chromosome Phylogenetic tree. This led to considerable confusion. In 2002, the major research groups came together and formed the Y-Chromosome Consortium (YCC). They published a joint paper that created a single new tree that all agreed to use. Later, a group of citizen scientists with an interest in population genetics and genetic genealogy formed a working group to create an amateur tree aiming at being above all timely. The table below brings together all of these works at the point of the landmark 2002 YCC Tree. This allows a researcher reviewing older published literature to quickly move between nomenclatures.
| YCC 2002/2008 (Shorthand) | (α) | (β) | (γ) | (δ) | (ε) | (ζ) | (η) | YCC 2002 (Longhand) | YCC 2005 (Longhand) | YCC 2008 (Longhand) | YCC 2010r (Longhand) | ISOGG 2006 | ISOGG 2007 | ISOGG 2008 | ISOGG 2009 | ISOGG 2010 | ISOGG 2011 | ISOGG 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E-P29 | 21 | III | 3A | 13 | Eu3 | H2 | B | E* | E | E | E | E | E | E | E | E | E | E |
| E-M33 | 21 | III | 3A | 13 | Eu3 | H2 | B | E1* | E1 | E1a | E1a | E1 | E1 | E1a | E1a | E1a | E1a | E1a |
| E-M44 | 21 | III | 3A | 13 | Eu3 | H2 | B | E1a | E1a | E1a1 | E1a1 | E1a | E1a | E1a1 | E1a1 | E1a1 | E1a1 | E1a1 |
| E-M75 | 21 | III | 3A | 13 | Eu3 | H2 | B | E2a | E2 | E2 | E2 | E2 | E2 | E2 | E2 | E2 | E2 | E2 |
| E-M54 | 21 | III | 3A | 13 | Eu3 | H2 | B | E2b | E2b | E2b | E2b1 | - | - | - | - | - | - | - |
| E-P2 | 25 | III | 4 | 14 | Eu3 | H2 | B | E3* | E3 | E1b | E1b1 | E3 | E3 | E1b1 | E1b1 | E1b1 | E1b1 | E1b1 |
| E-M2 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a* | E3a | E1b1 | E1b1a | E3a | E3a | E1b1a | E1b1a | E1b1a | E1b1a1 | E1b1a1 |
| E-M58 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a1 | E3a1 | E1b1a1 | E1b1a1 | E3a1 | E3a1 | E1b1a1 | E1b1a1 | E1b1a1 | E1b1a1a1a | E1b1a1a1a |
| E-M116.2 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a2 | E3a2 | E1b1a2 | E1b1a2 | E3a2 | E3a2 | E1b1a2 | E1b1a2 | E1ba12 | removed | removed |
| E-M149 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a3 | E3a3 | E1b1a3 | E1b1a3 | E3a3 | E3a3 | E1b1a3 | E1b1a3 | E1b1a3 | E1b1a1a1c | E1b1a1a1c |
| E-M154 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a4 | E3a4 | E1b1a4 | E1b1a4 | E3a4 | E3a4 | E1b1a4 | E1b1a4 | E1b1a4 | E1b1a1a1g1c | E1b1a1a1g1c |
| E-M155 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a5 | E3a5 | E1b1a5 | E1b1a5 | E3a5 | E3a5 | E1b1a5 | E1b1a5 | E1b1a5 | E1b1a1a1d | E1b1a1a1d |
| E-M10 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a6 | E3a6 | E1b1a6 | E1b1a6 | E3a6 | E3a6 | E1b1a6 | E1b1a6 | E1b1a6 | E1b1a1a1e | E1b1a1a1e |
| E-M35 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b* | E3b | E1b1b1 | E1b1b1 | E3b1 | E3b1 | E1b1b1 | E1b1b1 | E1b1b1 | removed | removed |
| E-M78 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b1* | E3b1 | E1b1b1a | E1b1b1a1 | E3b1a | E3b1a | E1b1b1a | E1b1b1a | E1b1b1a | E1b1b1a1 | E1b1b1a1 |
| E-M148 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b1a | E3b1a | E1b1b1a3a | E1b1b1a1c1 | E3b1a3a | E3b1a3a | E1b1b1a3a | E1b1b1a3a | E1b1b1a3a | E1b1b1a1c1 | E1b1b1a1c1 |
| E-M81 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b2* | E3b2 | E1b1b1b | E1b1b1b1 | E3b1b | E3b1b | E1b1b1b | E1b1b1b | E1b1b1b | E1b1b1b1 | E1b1b1b1a |
| E-M107 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b2a | E3b2a | E1b1b1b1 | E1b1b1b1a | E3b1b1 | E3b1b1 | E1b1b1b1 | E1b1b1b1 | E1b1b1b1 | E1b1b1b1a | E1b1b1b1a1 |
| E-M165 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b2b | E3b2b | E1b1b1b2 | E1b1b1b1b1 | E3b1b2 | E3b1b2 | E1b1b1b2a | E1b1b1b2a | E1b1b1b2a | E1b1b1b2a | E1b1b1b1a2a |
| E-M123 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b3* | E3b3 | E1b1b1c | E1b1b1c | E3b1c | E3b1c | E1b1b1c | E1b1b1c | E1b1b1c | E1b1b1c | E1b1b1b2a |
| E-M34 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b3a* | E3b3a | E1b1b1c1 | E1b1b1c1 | E3b1c1 | E3b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1b2a1 |
| E-M136 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3ba1 | E3b3a1 | E1b1b1c1a | E1b1b1c1a1 | E3b1c1a | E3b1c1a | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1b2a1a1 |
Scotland's DNA also found that more than 1% of all Scotsmen are direct descendants of the Berber and Tuareg tribesmen of the Sahara, a lineage which is around 5600 years old. Royal Stewart DNA was confirmed in 15% of male participants with the Stewart surname. They are directly descended from the royal line of kings. Source: https://www.bbc.com/news/uk-scotland-17740638.
Research publications
The following research teams per their publications were represented in the creation of the YCC tree.
Tree
This phylogenetic tree of haplogroup subclades is based on the Y-Chromosome Consortium (YCC) 2008 Tree,[59] the ISOGG Y-DNA Haplogroup E Tree,[7] and subsequent published research.
- E1b1a1 (DYS271/M2/SY81, M291, P1/PN1, P189, P293, V43, V95, Z1101, Z1107, Z1116, Z1120, Z1122, Z1123, Z1124, Z1125, Z1127, Z1130, Z1133) [Note 8]
- E1b1a1a (L576)
- E1b1a1a1 (L86.1, L88.3, M180/P88, PAGES00066, P182, Z1111, Z1112)
- E1b1a1a1a (M58, PAGES00027)
- E1b1a1a1b (M116.2)
- E1b1a1a1c (M149)
- E1b1a1a1d (M155)
- E1b1a1a1e (M10, M66, M156, M195)
- E1b1a1a1f (L485)
- E1b1a1a1f1 (L514)
- E1b1a1a1f1a (M191/P86, P253/U247, U186, Z1712, rs9786041)
- E1b1a1a1f1a1 (P252/U174)
- E1b1a1a1f1a1a (P9.2)
- E1b1a1a1f1a1b (P115)
- E1b1a1a1f1a1c (P116)
- E1b1a1a1f1a1c1 (P113)
- E1b1a1a1f1a1d (Z1704)
- (L372)
- E1b1a1a1f1a1 (P252/U174)
- E1b1a1a1f1b (L515, L516, L517, M263.2)
- E1b1a1a1f1b1 (Z1893)
- (Z1894)
- E1b1a1a1f1b1 (Z1893)
- E1b1a1a1f1a (M191/P86, P253/U247, U186, Z1712, rs9786041)
- E1b1a1a1f1 (L514)
- E1b1a1a1g (U175)
- E1b1a1a1g1 (L220.3, L652, P277, P278.1, U209, rs7067329, rs7474403, rs7893016)
- E1b1a1a1g1a (U290)
- E1b1a1a1g1a1 (U181)
- E1b1a1a1g1a1a (L97)
- E1b1a1a1g1a2 (Z1725)
- E1b1a1a1g1a1 (U181)
- E1b1a1a1g1b (P59)
- E1b1a1a1g1c (M154)
- E1b1a1a1g1d (V39)
- E1b1a1a1g1a (U290)
- E1b1a1a1g1 (L220.3, L652, P277, P278.1, U209, rs7067329, rs7474403, rs7893016)
- E1b1a1a1h (P268, P269)
- E1b1a1a1 (L86.1, L88.3, M180/P88, PAGES00066, P182, Z1111, Z1112)
- E1b1a1a (L576)
- E1b1a1 (DYS271/M2/SY81, M291, P1/PN1, P189, P293, V43, V95, Z1101, Z1107, Z1116, Z1120, Z1122, Z1123, Z1124, Z1125, Z1127, Z1130, Z1133) [Note 8]
| Phylogenetic tree of human Y-chromosome DNA haplogroups [χ 1][χ 2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-chromosomal Adam" | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| G | HIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | H | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| I | J | LT [χ 5] | K2 [χ 6] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| L | T | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| K-M2313 [χ 10] | K2b1 [χ 11] | P [χ 12] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NO | S [χ 13] | M [χ 14] | P1 | P2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N | O | Q | R | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
See also
| Wikiquote has quotations related to: Haplogroup E-M2 |
Genetics
- Genetic genealogy
- Haplogroup D
- Haplogroup DE
- Haplogroup
- Haplotype
- Human Y-chromosome DNA haplogroup
- Molecular phylogenetics
- Paragroup
- Subclade
- Y-chromosome haplogroups in populations of the world
- Y-DNA haplogroups by ethnic group
- Y-DNA haplogroups in populations of Sub-Saharan Africa
Notes
- All were positive for U175.
- The publication refers to E-V38 as H22.
- All E-M329, no E-M2 in this Eritrean data set
- All E-M329, no E-M2 in this Ethiopian data set
- E-M2 is approximately 7.7–7.9% of total US male population.
- The publication transposes M116.2 with M116.1 in Table 1.
- The YCAII STR marker value of 19–19 is also usually indicative of U175.
- DYS271/M2/SY81, P1/PN1, P189, P293, and M291 appear to form E1b1a1*. L576 forms a subclade immediately after the previously mentioned SNPs. L576 gave rise to a deeper subclade of M180/P88, P182, L88.3, L86, and PAGES0006. From this subclade, all the major subclades (i.e. E-U175 and E-L485) of E1b1a evolved. The exact position of V43 and V95 within these three subclades and E1b1a1a1b (M116.2), E1b1a1a1c (M149), and E1b1a1a1d (M155) remains uncertain.
References
- D'Atanasio E, Trombetta B, Bonito M, Finocchio A, Di Vito G, Seghizzi M; et al. (2018). "The peopling of the last Green Sahara revealed by high-coverage resequencing of trans-Saharan patrilineages". Genome Biol. 19 (1): 20. doi:10.1186/s13059-018-1393-5. PMC 5809971. PMID 29433568.CS1 maint: multiple names: authors list (link)
- "E-M2 YTree".
- Trombetta, Beniamino; D’Atanasio, Eugenia; Massaia, Andrea; Ippoliti, Marco; Coppa, Alfredo; Candilio, Francesca; Coia, Valentina; Russo, Gianluca; Dugoujon, Jean-Michel; Moral, Pedro; Akar, Nejat; Sellitto, Daniele; Valesini, Guido; Novelletto, Andrea; Scozzari, Rosaria; Cruciani, Fulvio (2015). "Phylogeographic Refinement and Large Scale Genotyping of Human Y Chromosome Haplogroup E Provide New Insights into the Dispersal of Early Pastoralists in the African Continent". Genome Biology and Evolution. Genome Biol Evol. 7 (7): 1940–1950. doi:10.1093/gbe/evv118. PMC 4524485. PMID 26108492.
- Shriner, Daniel; Rotimi, Charles (2018). "Whole-Genome-Sequence-Based Haplotypes Reveal Single Origin of the Sickle Allele during the Holocene Wet Phase". American Journal of Human Genetics. Am J Hum Genet. 102 (4): 547–556. doi:10.1016/j.ajhg.2018.02.003. PMC 5985360. PMID 29526279.
- Trombetta B, Cruciani F, Sellitto D, Scozzari R (January 2011). MacAulay V (ed.). "A new topology of the human Y chromosome haplogroup E1b1 (E-P2) revealed through the use of newly characterized binary polymorphisms". PLOS ONE. 6 (1): e16073. Bibcode:2011PLoSO...616073T. doi:10.1371/journal.pone.0016073. PMC 3017091. PMID 21253605.
- "E-V43 YTree".
- International Society of Genetic Genealogy (3 February 2010). "Y-DNA Haplogroup E and its Subclades – 2010". Retrieved 17 December 2010.
- Adams, Jonathan. "Africa During the Last 150,000 Years". Archived from the original on 1 May 2006. Retrieved 26 January 2011.
- Montano V, Ferri G, Marcari V, Batini C, Anyaele O, Destro-Bisol G, Comas D (July 2011). "The Bantu expansion revisited: a new analysis of Y chromosome variation in Central Western Africa". Molecular Ecology. 20 (13): 2693–708. doi:10.1111/j.1365-294X.2011.05130.x. PMID 21627702. S2CID 9951365.
- Rosa A, Ornelas C, Jobling MA, Brehm A, Villems R (July 2007). "Y-chromosomal diversity in the population of Guinea-Bissau: a multiethnic perspective". BMC Evolutionary Biology. 7: 124. doi:10.1186/1471-2148-7-124. PMC 1976131. PMID 17662131.
- Wood ET, Stover DA, Ehret C, Destro-Bisol G, Spedini G, McLeod H, Louie L, Bamshad M, Strassmann BI, Soodyall H, Hammer MF (July 2005). "Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes". European Journal of Human Genetics. 13 (7): 867–76. doi:10.1038/sj.ejhg.5201408. PMID 15856073.
- Underhill PA, Passarino G, Lin AA, Shen P, Mirazón Lahr M, Foley RA, Oefner PJ, Cavalli-Sforza LL (January 2001). "The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations". Annals of Human Genetics. 65 (Pt 1): 43–62. doi:10.1046/j.1469-1809.2001.6510043.x. PMID 11415522. S2CID 9441236.
- Luis JR, Rowold DJ, Regueiro M, Caeiro B, Cinnioğlu C, Roseman C, Underhill PA, Cavalli-Sforza LL, Herrera RJ (March 2004). "The Levant versus the Horn of Africa: evidence for bidirectional corridors of human migrations". American Journal of Human Genetics. 74 (3): 532–44. doi:10.1086/382286. PMC 1182266. PMID 14973781.
- Cruciani F, Santolamazza P, Shen P, Macaulay V, Moral P, Olckers A, Modiano D, Holmes S, Destro-Bisol G, Coia V, Wallace DC, Oefner PJ, Torroni A, Cavalli-Sforza LL, Scozzari R, Underhill PA (May 2002). "A back migration from Asia to sub-Saharan Africa is supported by high-resolution analysis of human Y-chromosome haplotypes". American Journal of Human Genetics. 70 (5): 1197–214. doi:10.1086/340257. PMC 447595. PMID 11910562.
- International HapMap Consortium (October 2005). "A haplotype map of the human genome". Nature. 437 (7063): 1299–320. Bibcode:2005Natur.437.1299T. doi:10.1038/nature04226. PMC 1880871. PMID 16255080.
- Semino O, Santachiara-Benerecetti AS, Falaschi F, Cavalli-Sforza LL, Underhill PA (January 2002). "Ethiopians and Khoisan share the deepest clades of the human Y-chromosome phylogeny". American Journal of Human Genetics. 70 (1): 265–8. doi:10.1086/338306. PMC 384897. PMID 11719903.
- Pereira L, Cerný V, Cerezo M, Silva NM, Hájek M, Vasíková A, Kujanová M, Brdicka R, Salas A (August 2010). "Linking the sub-Saharan and West Eurasian gene pools: maternal and paternal heritage of the Tuareg nomads from the African Sahel". European Journal of Human Genetics. 18 (8): 915–23. doi:10.1038/ejhg.2010.21. PMC 2987384. PMID 20234393.
- Msaidie S, Ducourneau A, Boetsch G, Longepied G, Papa K, Allibert C, Yahaya AA, Chiaroni J, Mitchell MJ (January 2011). "Genetic diversity on the Comoros Islands shows early seafaring as major determinant of human biocultural evolution in the Western Indian Ocean". European Journal of Human Genetics. 19 (1): 89–94. doi:10.1038/ejhg.2010.128. PMC 3039498. PMID 20700146.
- Gonçalves R, Rosa A, Freitas A, Fernandes A, Kivisild T, Villems R, Brehm A (November 2003). "Y-chromosome lineages in Cabo Verde Islands witness the diverse geographic origin of its first male settlers". Human Genetics. 113 (6): 467–72. doi:10.1007/s00439-003-1007-4. hdl:10400.13/3047. PMID 12942365. S2CID 63381583.
- Tofanelli S, Bertoncini S, Castrì L, Luiselli D, Calafell F, Donati G, Paoli G (September 2009). "On the origins and admixture of Malagasy: new evidence from high-resolution analyses of paternal and maternal lineages". Molecular Biology and Evolution. 26 (9): 2109–24. doi:10.1093/molbev/msp120. PMID 19535740.
- Sanchez JJ, Hallenberg C, Børsting C, Hernandez A, Morling N (July 2005). "High frequencies of Y chromosome lineages characterized by E3b1, DYS19-11, DYS392-12 in Somali males". European Journal of Human Genetics. 13 (7): 856–66. doi:10.1038/sj.ejhg.5201390. PMID 15756297.
- Iacovacci G, D'Atanasio E, Marini O, Coppa A, Sellitto D, Trombetta B, Berti A, Cruciani F (March 2017). "Forensic data and microvariant sequence characterization of 27 Y-STR loci analyzed in four Eastern African countries". Forensic Science International. Genetics. 27: 123–131. doi:10.1016/j.fsigen.2016.12.015. PMID 28068531.
- Plaster et al. Y-DNA E subclades
- Plaster CA (2011-09-28). Variation in Y chromosome, mitochondrial DNA and labels of identity on Ethiopia. discovery.ucl.ac.uk (Doctoral). Retrieved 2018-06-27.
- Ottoni C, Larmuseau MH, Vanderheyden N, Martínez-Labarga C, Primativo G, Biondi G, Decorte R, Rickards O (May 2011). "Deep into the roots of the Libyan Tuareg: a genetic survey of their paternal heritage". American Journal of Physical Anthropology. 145 (1): 118–24. doi:10.1002/ajpa.21473. PMID 21312181.
- Robino C, Crobu F, Di Gaetano C, Bekada A, Benhamamouch S, Cerutti N, Piazza A, Inturri S, Torre C (May 2008). "Analysis of Y-chromosomal SNP haplogroups and STR haplotypes in an Algerian population sample". International Journal of Legal Medicine. 122 (3): 251–5. doi:10.1007/s00414-007-0203-5. PMID 17909833. S2CID 11556974.
- Bosch E, Calafell F, Comas D, Oefner PJ, Underhill PA, Bertranpetit J (April 2001). "High-resolution analysis of human Y-chromosome variation shows a sharp discontinuity and limited gene flow between northwestern Africa and the Iberian Peninsula". American Journal of Human Genetics. 68 (4): 1019–29. doi:10.1086/319521. PMC 1275654. PMID 11254456.
- Cruciani F, La Fratta R, Santolamazza P, Sellitto D, Pascone R, Moral P, Watson E, Guida V, Colomb EB, Zaharova B, Lavinha J, Vona G, Aman R, Cali F, Akar N, Richards M, Torroni A, Novelletto A, Scozzari R (May 2004). "Phylogeographic analysis of haplogroup E3b (E-M215) y chromosomes reveals multiple migratory events within and out of Africa". American Journal of Human Genetics. 74 (5): 1014–22. doi:10.1086/386294. PMC 1181964. PMID 15042509.
- Karafet TM, Zegura SL, Posukh O, Osipova L, Bergen A, Long J, Goldman D, Klitz W, Harihara S, de Knijff P, Wiebe V, Griffiths RC, Templeton AR, Hammer MF (March 1999). "Ancestral Asian source(s) of new world Y-chromosome founder haplotypes". American Journal of Human Genetics. 64 (3): 817–31. doi:10.1086/302282. PMC 1377800. PMID 10053017.
- Arredi B, Poloni ES, Paracchini S, Zerjal T, Fathallah DM, Makrelouf M, Pascali VL, Novelletto A, Tyler-Smith C (August 2004). "A predominantly neolithic origin for Y-chromosomal DNA variation in North Africa". American Journal of Human Genetics. 75 (2): 338–45. doi:10.1086/423147. PMC 1216069. PMID 15202071.
- Hassan HY, Underhill PA, Cavalli-Sforza LL, Ibrahim ME (November 2008). "Y-chromosome variation among Sudanese: restricted gene flow, concordance with language, geography, and history". American Journal of Physical Anthropology. 137 (3): 316–23. doi:10.1002/ajpa.20876. PMID 18618658.
- Mršić G, Gršković B, Vrdoljak A, Popović M, Valpotić I, Anđelinović Š, Stenzl V, Ehler E, Urban L, Lacković G, Underhill P, Primorac D (July 2012). Branka Grskovic, Andro Vrdoljak, Maja Popovic, Ivica Valpotic, Simun Andelinovic, Vlastimil Stenzl, Edvard Ehler, Ludvik Urban, Gordana Lackovic, Peter Underhill, Dragan Primorac. "Croatian national reference Y-STR haplotype database". Molecular Biology Reports. 39 (7): 7727–41. doi:10.1007/s11033-012-1610-3. PMID 22391654. S2CID 18011987.
- Capelli C, Redhead N, Romano V, Calì F, Lefranc G, Delague V, et al. (March 2006). "Population structure in the Mediterranean basin: a Y chromosome perspective". Annals of Human Genetics. 70 (Pt 2): 207–25. doi:10.1111/j.1529-8817.2005.00224.x. hdl:2108/37090. PMID 16626331. S2CID 25536759.
- Flores C, Maca-Meyer N, González AM, Oefner PJ, Shen P, Pérez JA, Rojas A, Larruga JM, Underhill PA (October 2004). "Reduced genetic structure of the Iberian peninsula revealed by Y-chromosome analysis: implications for population demography". European Journal of Human Genetics. 12 (10): 855–63. doi:10.1038/sj.ejhg.5201225. PMID 15280900.
- Adams SM, Bosch E, Balaresque PL, Ballereau SJ, Lee AC, Arroyo E, López-Parra AM, Aler M, Grifo MS, Brion M, Carracedo A, Lavinha J, Martínez-Jarreta B, Quintana-Murci L, Picornell A, Ramon M, Skorecki K, Behar DM, Calafell F, Jobling MA (December 2008). "The genetic legacy of religious diversity and intolerance: paternal lineages of Christians, Jews, and Muslims in the Iberian Peninsula". American Journal of Human Genetics. 83 (6): 725–36. doi:10.1016/j.ajhg.2008.11.007. PMC 2668061. PMID 19061982.
- Abu-Amero KK, Hellani A, González AM, Larruga JM, Cabrera VM, Underhill PA (September 2009). "Saudi Arabian Y-Chromosome diversity and its relationship with nearby regions". BMC Genetics. 10: 59. doi:10.1186/1471-2156-10-59. PMC 2759955. PMID 19772609.
- Cadenas AM, Zhivotovsky LA, Cavalli-Sforza LL, Underhill PA, Herrera RJ (March 2008). "Y-chromosome diversity characterizes the Gulf of Oman". European Journal of Human Genetics. 16 (3): 374–86. doi:10.1038/sj.ejhg.5201934. PMID 17928816.
- Regueiro M, Cadenas AM, Gayden T, Underhill PA, Herrera RJ (2006). "Iran: tricontinental nexus for Y-chromosome driven migration". Human Heredity. 61 (3): 132–43. doi:10.1159/000093774. PMID 16770078. S2CID 7017701.
- Al-Zahery N, Semino O, Benuzzi G, Magri C, Passarino G, Torroni A, Santachiara-Benerecetti AS (September 2003). "Y-chromosome and mtDNA polymorphisms in Iraq, a crossroad of the early human dispersal and of post-Neolithic migrations". Molecular Phylogenetics and Evolution. 28 (3): 458–72. doi:10.1016/S1055-7903(03)00039-3. PMID 12927131.
- Firasat S, Khaliq S, Mohyuddin A, Papaioannou M, Tyler-Smith C, Underhill PA, Ayub Q (January 2007). "Y-chromosomal evidence for a limited Greek contribution to the Pathan population of Pakistan". European Journal of Human Genetics. 15 (1): 121–6. doi:10.1038/sj.ejhg.5201726. PMC 2588664. PMID 17047675.
- Cinnioğlu C, King R, Kivisild T, Kalfoğlu E, Atasoy S, Cavalleri GL, Lillie AS, Roseman CC, Lin AA, Prince K, Oefner PJ, Shen P, Semino O, Cavalli-Sforza LL, Underhill PA (January 2004). "Excavating Y-chromosome haplotype strata in Anatolia". Human Genetics. 114 (2): 127–48. doi:10.1007/s00439-003-1031-4. PMID 14586639. S2CID 10763736.
- Sims LM, Garvey D, Ballantyne J (January 2007). "Sub-populations within the major European and African derived haplogroups R1b3 and E3a are differentiated by previously phylogenetically undefined Y-SNPs". Human Mutation. 28 (1): 97. doi:10.1002/humu.9469. PMID 17154278. S2CID 34556775.
- Mendizabal I, Sandoval K, Berniell-Lee G, Calafell F, Salas A, Martínez-Fuentes A, Comas D (July 2008). "Genetic origin, admixture, and asymmetry in maternal and paternal human lineages in Cuba". BMC Evolutionary Biology. 8: 213. doi:10.1186/1471-2148-8-213. PMC 2492877. PMID 18644108.
- Bryc K, Velez C, Karafet T, Moreno-Estrada A, Reynolds A, Auton A, Hammer M, Bustamante CD, Ostrer H (May 2010). "Colloquium paper: genome-wide patterns of population structure and admixture among Hispanic/Latino populations". Proceedings of the National Academy of Sciences of the United States of America. 107 Suppl 2 (Supplement 2): 8954–61. Bibcode:2010PNAS..107.8954B. doi:10.1073/pnas.0914618107. PMC 3024022. PMID 20445096.
- Nuñez C, Baeta M, Sosa C, Casalod Y, Ge J, Budowle B, Martínez-Jarreta B (December 2010). "Reconstructing the population history of Nicaragua by means of mtDNA, Y-chromosome STRs, and autosomal STR markers". American Journal of Physical Anthropology. 143 (4): 591–600. doi:10.1002/ajpa.21355. PMID 20721944.
- Rojas W, Parra MV, Campo O, Caro MA, Lopera JG, Arias W, Duque C, Naranjo A, García J, Vergara C, Lopera J, Hernandez E, Valencia A, Caicedo Y, Cuartas M, Gutiérrez J, López S, Ruiz-Linares A, Bedoya G (September 2010). "Genetic make up and structure of Colombian populations by means of uniparental and biparental DNA markers". American Journal of Physical Anthropology. 143 (1): 13–20. doi:10.1002/ajpa.21270. PMID 20734436.
- de Azevedo DA, da Silva LA, Gusmão L, de Carvalho EF (December 2009). "Analysis of Y chromosome SNPs in Alagoas, Northeastern Brazil". Forensic Science International: Genetics Supplement Series. 2 (1): 421–422. doi:10.1016/j.fsigss.2009.08.166.
- Nascimento E, Cerqueira E, Azevedo E, Freitas V, Azevedo D (December 2009). "The Africa male lineages of Bahia's people—Northeast Brazil: A preliminary SNPs study". Forensic Science International: Genetics Supplement Series. 2 (1): 349–350. doi:10.1016/j.fsigss.2009.07.010.
- Tanya M Simms 2011, The Peopling of the Bahamas: A Phylogeographical Perspective pg. 194
- Hünemeier T, Carvalho C, Marrero AR, Salzano FM, Pena SD, Bortolini MC (June 2007). "Niger-Congo speaking populations and the formation of the Brazilian gene pool: mtDNA and Y-chromosome data". American Journal of Physical Anthropology. 133 (2): 854–67. doi:10.1002/ajpa.20604. PMID 17427922.
- de Filippo C, Barbieri C, Whitten M, Mpoloka SW, Gunnarsdóttir ED, Bostoen K, Nyambe T, Beyer K, Schreiber H, de Knijff P, Luiselli D, Stoneking M, Pakendorf B (March 2011). "Y-chromosomal variation in sub-Saharan Africa: insights into the history of Niger-Congo groups". Molecular Biology and Evolution. 28 (3): 1255–69. doi:10.1093/molbev/msq312. PMC 3561512. PMID 21109585.
- Veeramah KR, Connell BA, Ansari Pour N, Powell A, Plaster CA, Zeitlyn D, Mendell NR, Weale ME, Bradman N, Thomas MG (March 2010). "Little genetic differentiation as assessed by uniparental markers in the presence of substantial language variation in peoples of the Cross River region of Nigeria". BMC Evolutionary Biology. 10: 92. doi:10.1186/1471-2148-10-92. PMC 2867817. PMID 20356404.
- Naidoo T, Schlebusch CM, Makkan H, Patel P, Mahabeer R, Erasmus JC, Soodyall H (September 2010). "Development of a single base extension method to resolve Y chromosome haplogroups in sub-Saharan African populations". Investigative Genetics. 1 (1): 6. doi:10.1186/2041-2223-1-6. PMC 2988483. PMID 21092339.
- Abecasis GR, Altshuler D, Auton A, Brooks LD, Durbin RM, Gibbs RA, Hurles ME, McVean GA (October 2010). "A map of human genome variation from population-scale sequencing". Nature. 467 (7319): 1061–73. Bibcode:2010Natur.467.1061T. doi:10.1038/nature09534. PMC 3042601. PMID 20981092.
- Reynolds D, Squecco A. "Y-Chromosome Genome Comparison". Retrieved 1 August 2011.
- Brucato N, Cassar O, Tonasso L, Tortevoye P, Migot-Nabias F, Plancoulaine S, Guitard E, Larrouy G, Gessain A, Dugoujon JM (October 2010). "The imprint of the Slave Trade in an African American population: mitochondrial DNA, Y chromosome and HTLV-1 analysis in the Noir Marron of French Guiana". BMC Evolutionary Biology. 10: 314. doi:10.1186/1471-2148-10-314. PMC 2973943. PMID 20958967.
- Brito P, Carvalho M, Gomes V, Melo MM, Bogas V, Balsa F, et al. (December 2011). "Y-SNP analysis in an Angola population". Forensic Science International: Genetics Supplement Series. 3 (1): e369–e370. doi:10.1016/j.fsigss.2011.09.046.
- Underhill PA, Shen P, Lin AA, Jin L, Passarino G, Yang WH, Kauffman E, Bonné-Tamir B, Bertranpetit J, Francalacci P, Ibrahim M, Jenkins T, Kidd JR, Mehdi SQ, Seielstad MT, Wells RS, Piazza A, Davis RW, Feldman MW, Cavalli-Sforza LL, Oefner PJ (November 2000). "Y chromosome sequence variation and the history of human populations". Nature Genetics. 26 (3): 358–61. doi:10.1038/81685. PMID 11062480. S2CID 12893406.
- Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF (May 2008). "New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree". Genome Research. 18 (5): 830–8. doi:10.1101/gr.7172008. PMC 2336805. PMID 18385274.
