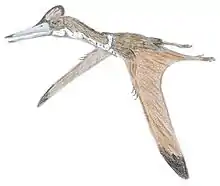
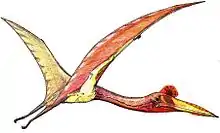
Hatzegopteryx
Hatzegopteryx ("Hațeg basin wing") is a genus of azhdarchid pterosaur found in the late Maastrichtian deposits of the Densuş Ciula Formation, an outcropping in Transylvania, Romania. It is known only from the type species, Hatzegopteryx thambema, named by Buffetaut et al. in 2002 based on parts of the skull and humerus. Additional specimens, including a neck vertebra, were later placed in the genus, representing a range of sizes. The largest of these remains indicate it was among the biggest pterosaurs, with an estimated wingspan of 10 to 12 metres (33 to 39 ft).
| Hatzegopteryx | |
|---|---|
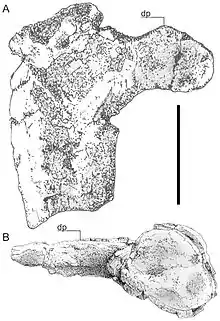 | |
| Left humerus of the holotype specimen in ventral (A) and distal (B) view | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Order: | †Pterosauria |
| Suborder: | †Pterodactyloidea |
| Family: | †Azhdarchidae |
| Genus: | †Hatzegopteryx Buffetaut et al., 2002 |
| Type species | |
| †Hatzegopteryx thambema Buffetaut et al., 2002 | |
Unusually among giant azhdarchids, Hatzegopteryx had a very wide skull bearing large muscular attachments; bones with a spongy internal texture instead of hollow; and a short, robust, and heavily muscled neck measuring 1.5 metres (5 ft) long, which was about half the length of other azhdarchids with comparable wingspans, and was capable of withstanding strong bending forces. Hatzegopteryx inhabited Hațeg Island, an island situated in the Cretaceous subtropics within the prehistoric Tethys Sea. In the absence of large theropods, Hatzegopteryx was likely the apex predator of Hațeg Island, tackling proportionally larger prey (including dwarf titanosaurs and iguanodontians) than other azhdarchids.
Discovery and naming

The first remains of Hatzegopteryx were found from the upper part of the Middle Densuş Ciula Formation of Vălioara, northwestern Hațeg Basin, Transylvania, western Romania, which has been dated to the late Maastrichtian stage of the Late Cretaceous Period, around 66 million years ago. The holotype of Hatzegopteryx, FGGUB R 1083A, consists of two fragments from the back of the skull and the damaged proximal part of a left humerus.[1] One of these fragments, namely the occipital region, was initially referred to a theropod dinosaur when it was first announced in 1991.[2][3] A 38.5 cm (15.2 in) long middle section of a femur found nearby, FGGUB R1625, may also belong to Hatzegopteryx.[4] FGGUB R1625 would have belonged to a smaller individual of Hatzegopteryx (assuming it pertains to the genus), with a 5 to 6 m (16 to 20 ft) wingspan. Additional reported specimens from the locality include an unpublished mandible, also from a large individual.[5][6]
Hatzegopteryx was named in 2002 by French paleontologist Eric Buffetaut, and Romanian paleontologists Dan Grigorescu and Zoltan Csiki. The generic name is derived from the Hatzeg (or Hațeg) basin of Transylvania, where the bones were found, and from Greek pteryx (ἡ πτέρυξ, -υγος (also ἡ πτερύξ, -ῦγος), or 'wing'. The specific name thambema is derived from the Greek for 'terror, monster' (τό θάμβημα, -ήματος[7]), in reference to its huge size.[1]
New specimens of Hatzegopteryx have since been recovered from other localities. In the Sânpetru Formation from the locality of Vadu, Sântămăria-Orlea, a medium-sized scapulocoracoid was found, which probably pertained to an individual with a wingspan of 4.5 to 5 m (15 to 16 ft). From the Râpa Roşie locality of the Sebeş Formation, which is contemporary and adjacent to the Densuș Ciula Formation, a single large neck vertebra, the "RR specimen" or EME 215, was found.[5][6] Although the lack of overlapping elements prevents this specimen from being definitely referred to Hatzegopteryx thambena, its distinctive internal bone structure, as well as the lack of evidence for a second giant azhdarchid in the area, warrant its referral to at least H. sp.[8]
Description
Skull
The skull of Hatzegopteryx was giant, with an initially estimated length of 3 metres (9.8 ft) based on comparisons with Nyctosaurus and Anhanguera, making it one of the largest skulls among non-marine animals. The skull was broadened in the rear, being 0.5 metres (1 ft 8 in) wide across the quadrate bones.[4] While most pterosaur skulls are composed of gracile plates and struts, in Hatzegopteryx the skull bones are stout and robust, with large ridges indicating strong muscular attachments.[1] In 2018, Mátyás Vremir concluded that Hatzegopteryx likely had a shorter and broader skull, the length of which he estimated at 1.6 metres.[9]
The massive jaw bore a distinctive groove at its point of articulation (also seen in some other pterosaurs, including Pteranodon) that would have allowed the animal to achieve a very wide gape.[1] Unpublished remains attributed to Hatzegopteryx suggest that it had a proportionally short, deep beak, grouping with the "blunt-beaked" azhdarchids rather than the "slender-beaked" azhdarchids (including Quetzalcoatlus sp.).[10]
Cervical vertebrae

A large neck vertebra attributed to Hatzegopteryx is short and unusually robust. The preserved portion measures 240 millimetres (9.4 in) long, with the entire vertebra likely measuring 300 millimetres (12 in) long in life.[5] Pterosaurs had nine neck vertebrae;[11] regression indicates that the third to seventh cervical vertebrae would have collectively measured 1.508 metres (4 ft 11.4 in) in length, with the longest vertebra - the fifth - only measuring approximately 400 mm (16 in) long. Meanwhile, the same vertebrae in the similarly-giant Arambourgiania measured 2.652 metres (8 ft 8.4 in); this indicates that the neck of Hatzegopteryx is about 50–60% the length of what would be expected for a giant azhdarchid of its size.[8]
The bottom surface of the neck vertebra was also unusually thick, at 4 to 6 millimetres (0.16 to 0.24 in). For most other giant azhdarchids, including Arambourgiania, this surface is less than 2.6 millimetres (0.10 in) thick. Although the neural spine of the vertebra is not completely preserved, the width of the preserved portion suggests that it was relatively tall and robust relative to those of other pterosaurs. Other aspects of the vertebra converge upon the seventh neck vertebra of the smaller Azhdarcho most closely: the articulating sockets (cotyles) are much shallower than the neural arches, and are four times as wide as they are tall; a process on the bottom of the vertebrae, known as a hypapophysis, is present; the processes at the front of the vertebrae, the prezygapophyses, are splayed; and the vertebra has a tapered "waist" in the middle of the centrum.[8] Although initially identified as a third neck vertebra,[5] these traits supports the identification of the vertebra as coming from the rear of the neck, more specifically as being the seventh vertebra.[8]
Size

The size of Hatzegopteryx was initially estimated by comparing the 236 mm (9.3 in) humerus fragment with that of Quetzalcoatlus northropi, which has a 544 mm (21.4 in)-long humerus. Observing that the Hatzegopteryx fragment presented less than half of the original bone, Buffetaut et al. established that it could possibly have been "slightly longer" than that of Quetzalcoatlus. The wingspan of the latter had been estimated at 11 to 12 metres (36 to 39 ft) in 1981; earlier estimates had strongly exceeded this at 15 to 20 metres (49 to 66 ft). They concluded that an estimate of a 12 metres (39 ft) wing span for Hatzegopteryx was conservative, "provided that its humerus was longer than that of Q. northropi".[1][4] In 2010, Mark Witton and Michael Habib concluded that Hatzegopteryx was probably no larger than Q. northropi in wingspan; the initial conclusions did not account for distortion of the bone. The latter is generally estimated at 10–11 metres (33–36 ft) in length.[12]
It has been suggested, on the basis of the wide and robust neck vertebra referred to Hatzegopteryx, that the entire vertebral column of the animal was similarly expanded, increasing its overall size.[5] However, this is likely not true, since the neck vertebrae of large pterodactyloids generally tend to be wider and larger than the rest of the vertebrae. Although estimates of pterosaur size based on vertebrae alone are not particularly reliable, the size of this vertebra is consistent with an animal that measured 10 to 12 metres (33 to 39 ft) in wingspan.[8]
Classification
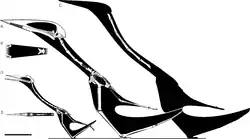
Similarities between the humerus of Hatzegopteryx and Quetzalcoatlus northropi have been noted; both have a long, smooth deltopectoral crest, and a thickened humeral head. These were initially the basis of the taxon's referral to the clade Azhdarchidae,[1] but they are also similar enough to be a basis for the synonymy of Hatzegopteryx and Quetzalcoatlus. However, this is likely due to the relatively non-diagnostic nature of the humerus in giant azhdarchid taxonomy, and the lack of a detailed description for the elements of Q. northropi.[13] However, the neck and jaw anatomy of Hatzegopteryx is quite clearly distinct from the smaller Q. sp., which warrants the retention of Hatzegopteryx as a taxon separate from Quetzalcoatlus.[1][8][14]
The neck vertebra referred to Hatzegopteryx sp. contains a number of traits that allow for it to be definitely identified as that of an azhdarchid. The centrum is relatively low, the zygapophyses are large and flattened, and the preserved portions of the neural spine indicate that it is bifid, or split into two.[8]
Paleobiology
Bone structure
While the skull of Hatzegopteryx was unusually large and robust, its wing bones are comparable to those of other flying pterosaurs, indicating that it was not flightless. Buffetaut et al. suggested that, in order to fly, the skull weight of Hatzegopteryx must have been reduced in some way. The necessary weight reduction may have accomplished by the internal structure of the skull bones, which were full of small pits and hollows (alveoli) up to 10 millimetres (0.39 in) long, separated by a matrix of thin bony struts (trabeculae). The wing bones also bear a similar internal structure. This unusual construction differs from that of other pterosaurs, and resembles more closely the structure of expanded polystyrene (used to manufacture Styrofoam). This would have made the skull sturdy and stress-resistant, but also lightweight, enabling the animal to fly.[1] A similar internal structure is also seen in the cervical vertebra referred to Hatzegopteryx.[8]
Neck biomechanics

As a consequence of its robust, thick-walled vertebrae, the neck of Hatzegopteryx was much stronger than that of Arambourgiania. This can be quantified using relative failure force, which is the bone failure force of a vertebra divided by the body weight of the pterosaur that it belongs, estimated at 180 to 250 kg (400 to 550 lb) for Arambourgiania and Hatzegopteryx. While Arambourgiania's neck vertebrae fail at about half of its body weight, the posterior neck vertebrae of Hatzegopteryx can withstand anywhere between five to ten body weights, depending on the loading of the bone. Even the hypothetically longer anterior neck vertebrae of Hatzegopteryx would be able to withstand four to seven body weights.[8]

Although the centrum of Hatzegopteryx is much more robust than Arambourgiania, their ratios of bone radius to bone thickness (R/t)[12] are roughly the same (9.45 for Hatzegopteryx and 9.9 for Arambourgiania). This may represent a compromise between increasing bending strength and buckling strength; higher R/t ratios lead to improved bending strength, but weaker buckling strength. To compensate for this, Hatzegopteryx shows a number of other adaptations to improve buckling strength, namely the distinctive internal structures of the bones and the large articular joints of the vertebrae, the latter of which helps to distribute stress.[8]
In order to support the robust head, the neck of Hatzegopteryx was likely strongly muscled. On the occipital bones, the nuchal lines, which serve as muscular attachments, are very well-developed and bear prominent scars. These conceivably supported the transversospinalis muscles, which aid in extension and flexion of the head and neck. Likewise, the opisthotic process, neural spines, and zygapophyses all appeared to have been large and robust (with the latter bearing many pits and edges that likely represent muscle scars), and the basioccipital tuberosities were long; these all serve as points of attachment for various muscles of the head and neck. Although not entirely unmuscled, the neck of Arambourgiania probably would not have been as extensively muscled as that of Hatzegopteryx.[8]
Paleoecology

Like all azhdarchid pterosaurs, Hatzegopteryx was probably a terrestrially foraging generalist predator.[15] It is significantly larger than any other terrestrial predator from Maastrichtian Europe; due to its large size in an environment otherwise dominated by island dwarf dinosaurs, with no large hypercarnivorous theropods in the region, it has been suggested that Hatzegopteryx played the role of an apex predator in the Haţeg Island ecosystem. The robust anatomy of Hatzegopteryx suggests that it may have tackled larger prey than other azhdarchids, including animals too large to swallow whole; similarly, some modern storks (particularly the marabou stork and the jabiru) are known to attack and kill large prey such as flamingoes, and occasionally children, with their beaks.[15] Meanwhile, other giant azhdarchids like Arambourgiania would probably have instead fed on small prey (up to the size of a human), including hatchling or small dinosaurs and eggs.[8] Another pterosaur, Thalassodromeus, has similarly been suggested to be raptorial.[16]

Apart from Hatzegopteryx, there are various other unusual denizens of the Haţeg Island ecosystem. Co-occurring pterosaurs included the small azhdarchid Eurazhdarcho, with a wingspan of 3 m (9.8 ft);[6] an unnamed, small-sized short-necked azhdarchid with a wingspan of 3.5 to 4 metres (11 to 13 ft); a somewhat larger and likewise unnamed azhdarchid, with a wingspan of 5 m (16 ft); and apparently small pteranodontids have been found as well.[17] The robust, flightless, and possibly herbivorous avialan[18] or dromaeosaurid[19] Balaur, which had two enlarged claws on each foot,[19] represents another highly specialized component of the fauna. The ecosystem contained a number of insular dwarfs, namely the titanosaurs Magyarosaurus[20] and Paludititan,[21] the hadrosaurid Telmatosaurus, and the iguanodontian Zalmoxes.[20] Along with the nodosaurid Struthiosaurus, various small, fragmentary maniraptorans were present: Bradycneme, Elopteryx, and Heptasteornis.[20] Crocodilian remains, belonging to the genera Allodaposuchus, Doratodon, and Acynodon have also been found.[22] Non-archosaurian components include the kogaionid multituberculate mammals Kogaionon, Barbatodon, Litovoi tholocephalos, and Hainina;[23][24] lizards such as the teiid Bicuspidon and the paramacellodid Becklesius; an unnamed madtsoiid snake; and the lissamphibians Albanerpeton, Eodiscoglossus, and Paradiscoglossus.[25]
During the Maastrichtian, southern Europe was an archipelago. The members of the Haţeg Island ecosystem lived on a landmass known as the Tisia–Dacia Block, of which the Haţeg Basin was a small part. This landmass was about 80,000 square kilometres (31,000 sq mi) in area, and was separated from other terrestrial terrains by stretches of deep ocean in all directions by 200 to 300 kilometres (120 to 190 mi).[20] Being located at 27°N,[26] the island was located farther south than the present-day latitude of 45°N; the climate was likely subtropical, with distinct dry and wet seasons, and an average temperature of about 25 °C (77 °F).[27] The environment consisted of various alluvial plains, wetlands, and rivers,[28] surrounded by woodlands dominated by ferns and angiosperms.[20] Paleosols indicate a relatively dry Cretaceous climate, with an annual precipitation of less than 1,000 mm (39 in).[27]
References
- Buffetaut, E.; Grigorescu, D.; Csiki, Z. (2002). "A new giant pterosaur with a robust skull from the latest Cretaceous of Romania". Naturwissenschaften. 89 (4): 180–184. Bibcode:2002NW.....89..180B. doi:10.1007/s00114-002-0307-1. PMID 12061403. S2CID 15423666.
- Weishampel, D.B.; Barrett, P.M.; Coria, R.A.; Le Loeuff, J.; Xu, X.; Zhao, X.; Sahni, A.; Gomani, E.M.P.; Noto, C.M. (2004). "Dinosaur Distribution". In Weishampel, D.B.; Dodson, P.; Osmolska, H. (eds.). The Dinosauria (2nd ed.). University of California Press. p. 593. ISBN 9780520254084.
- Witton, M.P. (2017). "New paper: when the short-necked, giant azhdarchid pterosaur Hatzegopteryx ruled Late Cretaceous Romania". Markwitton.com Blog.
- Buffetaut, E.; Grigorescu, D.; Csiki, Z. (2003). "Giant azhdarchid pterosaurs from the terminal Cretaceous of Transylvania (western Romania)". Geological Society, London, Special Publications. 217 (1): 91–104. Bibcode:2003GSLSP.217...91B. doi:10.1144/GSL.SP.2003.217.01.09. S2CID 140190857.
- Vremir, M.M. (2010). "New faunal elements from the Late Cretaceous (Maastrichtian) continental deposits of Sebeş area (Transylvania)" (PDF). Acta Musei Sabesiensis. 2: 635–684.
- Vremir, M.; Kellner, A.W.A.; Naish, D.; Dyke, G.J. (2013). "A New Azhdarchid Pterosaur from the Late Cretaceous of the Transylvanian Basin, Romania: Implications for Azhdarchid Diversity and Distribution". PLOS ONE. 8 (1): e54268. Bibcode:2013PLoSO...854268V. doi:10.1371/journal.pone.0054268. PMC 3559652. PMID 23382886.
- Liddell, H.G.; Scott, R. (1940). "θάμβ-ημα". Perseus Digital Library. A Greek-English Lexicon. Tufts University.
- Naish, D.; Witton, M.P. (2017). "Neck biomechanics indicate that giant Transylvanian azhdarchid pterosaurs were short-necked arch predators". PeerJ. 5: e2908. doi:10.7717/peerj.2908. PMC 5248582. PMID 28133577.
- Mátyás Vremir, Gareth Dyke, Zoltán Csikiá Sava, Dan Grigorescu & Eric Buffetaut, 2018, "Partial mandible of a giant pterosaur from the uppermost Cretaceous (Maastrichtian) of the Haţeg Basin, Romania", Lethaia
- Witton, M.; Brusatte, S.; Dyke, G.; Naish, D.; Norell, M.; Vremir, M. (2013). Pterosaur overlords of Transylvania: short-necked giant azhdarchids in Late Cretaceous Romania. The Annual Symposium of Vertebrate Paleontology and Comparative Anatomy. Edinburgh. Archived from the original on 2016-04-06. Retrieved 2017-01-18.
- Bennett, S.C. (2014). "A new specimen of the pterosaur Scaphognathus crassirostris, with comments on constraint of cervical vertebrae number in pterosaurs". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 271 (3): 327–348. doi:10.1127/0077-7749/2014/0392.
- Witton, M.P.; Habib, M.B. (2010). "On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness". PLOS ONE. 5 (11): e13982. Bibcode:2010PLoSO...513982W. doi:10.1371/journal.pone.0013982. PMC 2981443. PMID 21085624.
- Witton, M.P.; Martill, D.M.; Loveridge, R.F. (2010). Clipping the Wings of Giant Pterosaurs: Comments on Wingspan Estimations and Diversity. Proceedings of the Third International Symposium on Pterosaurs. Acta Geoscientica Sinica. 31. Beijing. pp. 79–81. Archived from the original on 2017-01-31. Retrieved 2017-01-18.
- Witton, M.P. (2016). "Quetzalcoatlus: the media concept vs the science". Markwitton.com Blog.
- Witton, M.P.; Naish, D. (2015). "Azhdarchid pterosaurs: water-trawling pelican mimics or "terrestrial stalkers"?". Acta Palaeontologica Polonica. 60 (3). doi:10.4202/app.00005.2013.
- Witton, M.P. (2013). Pterosaurs: Natural History, Evolution, Anatomy. Princeton: Princeton University Press. p. 243. ISBN 9780691150611.
- Vremir, M.; Witton, M.; Naish, D.; Dyke, G.; Brusatte, S.L.; Norell, M.; Totoianu, R. (2015). "A medium-sized robust-necked azhdarchid pterosaur (Pterodactyloidea: Azhdarchidae) from the Maastrichtian of Pui (Haţeg Basin, Transylvania, Romania)". American Museum Novitates. 3827 (3827): 1–16. doi:10.1206/3827.1. hdl:2246/6582. ISSN 0003-0082. S2CID 54068293.
- Cau, A.; Brougham, T.; Naish, D. (2015). "The phylogenetic affinities of the bizarre Late Cretaceous Romanian theropod Balaur bondoc (Dinosauria, Maniraptora): dromaeosaurid or flightless bird?". PeerJ. 3: e1032. doi:10.7717/peerj.1032. PMC 4476167. PMID 26157616.
- Csiki, Z.; Vremir, M.; Brusatte, S.L.; Norell, M.A. (2010). "An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania". Proceedings of the National Academy of Sciences of the United States of America. 107 (35): 15357–15361. Bibcode:2010PNAS..10715357C. doi:10.1073/pnas.1006970107. PMC 2932599. PMID 20805514.
- Benton, M.J.; Csiki, Z.; Grigorescu, D.; Redelstorff, R.; Sander, P.M.; Stein, K.; Weishampel, D.B. (2010). "Dinosaurs and the island rule: The dwarfed dinosaurs from Haţeg Island". Palaeogeography, Palaeoclimatology, Palaeoecology. 293 (3): 438–454. Bibcode:2010PPP...293..438B. doi:10.1016/j.palaeo.2010.01.026.
- Cziki, Z.; Codrea, V.; Jipa-Murzea, C.; Godefroit, P. (2010). "A partial titanosaur (Sauropoda, Dinosauria) skeleton from the Maastrichtian of Nălaţ-Vad, Haţeg Basin, Romania". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 258 (3): 297–324. doi:10.1127/0077-7749/2010/0098.
- Martin, J.E.; Csiki, Z.; Grigorescu, D.; Buffetaut, E. (2006). "Late Cretaceous crocodilian diversity in Haţeg Basin, Romania". Hantkeniana. 5 (5): 31–37.
- Csiki, Z.; Grigorescu, D. (2000). "Teeth of multituberculate mammals from the Late Cretaceous of Romania". Acta Palaeontologica Polonica. 45 (1): 85–90.
- Codrea, V.A.; Solomon, A.A.; Venzcel, M.; Smith, T. (2017). "Première identification d'une espèce de mammifère du Crétacé supérieur du bassin de Rusca Montană (Transylvanie, Roumanie)" [First mammal species identified from the Upper Cretaceous of the Rusca Montană Basin (Transylvania, Romania)]. Comptes Rendus Palevol. 16 (1): 27–38. doi:10.1016/j.crpv.2016.04.002.
- Folie, A.; Codrea, V. (2005). "New lissamphibians and squamates from the Maastrichtian of Haţeg Basin, Romania". Acta Palaeontologica Polonica. 50 (1): 57–71.
- Panaiotu, C.G.; Panaiotu, C.E. (2010). "Palaeomagnetism of the Upper Cretaceous Sânpetru Formation (Haţeg Basin, South Carpathians)". Palaeogeography, Palaeoclimatology, Palaeoecology. 293 (4): 343–352. Bibcode:2010PPP...293..343P. doi:10.1016/j.palaeo.2009.11.017.
- Therrien, F. (2005). "Palaeoenvironments of the latest Cretaceous (Maastrichtian) dinosaurs of Romania: insights from fluvial deposits and paleosols of the Transylvanian and Hateg basins". Palaeogeography, Palaeoclimatology, Palaeoecology. 218 (1): 15–56. Bibcode:2005PPP...218...15T. doi:10.1016/j.palaeo.2004.12.005.
- Therrien, F.; Zelenitsky, D.K.; Weishampel, D.B. (2009). "Palaeoenvironmental reconstruction of the Late Cretaceous Sânpetru Formation (Haţeg Basin, Romania) using paleosols and implications for the "disappearance" of dinosaurs". Palaeogeography, Palaeoclimatology, Palaeoecology. 272 (1): 37–52. Bibcode:2009PPP...272...37T. doi:10.1016/j.palaeo.2008.10.023.
External links
![]() Media related to Hatzegopteryx at Wikimedia Commons
Media related to Hatzegopteryx at Wikimedia Commons