Induan
The Induan is the first age of the Early Triassic epoch in the geologic timescale, or the lowest stage of the Lower Triassic series in chronostratigraphy. It spans the time between 251.902 Ma and 251.2 Ma (million years ago).[7] The Induan is sometimes divided into the Griesbachian and the Dienerian subages or substages.[8] The Induan is preceded by the Changhsingian (latest Permian) and is followed by the Olenekian.
| Induan | |
|---|---|
| 251.902 ± 0.024 – 251.2 Ma | |
| Chronology | |
Key events in the Triassic -255 — – -250 — – -245 — – -240 — – -235 — – -230 — – -225 — – -220 — – -215 — – -210 — – -205 — – -200 — An approximate timescale of key Triassic events. Axis scale: millions of years ago. (NOTE: The white links are for readability only. They are still clickable.) | |
| Etymology | |
| Name formality | Formal |
| Usage information | |
| Celestial body | Earth |
| Regional usage | Global (ICS) |
| Time scale(s) used | ICS Time Scale |
| Definition | |
| Chronological unit | Age |
| Stratigraphic unit | Stage |
| Time span formality | Formal |
| Lower boundary definition | FAD of the Conodont Hindeodus parvus |
| Lower boundary GSSP | Meishan, Zhejiang, China 31.0798°N 119.7058°E |
| GSSP ratified | 2001[5] |
| Upper boundary definition | Not formally defined |
| Upper boundary definition candidates | FAD of the Conodont Neospathodus waageni |
| Upper boundary GSSP candidate section(s) | Mud (Muth) village, Spiti valley, India[6] |
.jpg.webp)
The Induan is roughly coeval with the regional Feixianguanian stage of China.
Stratigraphic definitions

The Induan stage was introduced into scientific literature by Russian stratigraphers in 1956,[9] who divided the Scythian stage that was used by Western stratigraphers into the Induan and Olenekian stages. The Induan stage is named for the Indus region of India.[10] The Russian subdivision of the Lower Triassic then slowly replaced the one used in the West.
The base of the Induan stage (which is also the base of the Lower Triassic series, the base of the Triassic system and the base of the Mesozoic erathem) is defined as the place in the fossil record where the conodont species Hindeodus parvus first appears, or at the end of the negative δ18O anomaly after the big extinction event at the Permian-Triassic boundary. The global reference profile of the base of the Induan is situated in Meishan, Changxing County, China.[11]
The top of the Induan stage (the base of the Olenekian) is at the first appearance of ammonite species Meekoceras gracilitatis.
Though the Induan is an unusually short age at this point in the geologic timescale, its million years' extent still contains five ammonite biozones in the boreal domain and four ammonite biozones in the Tethyan domain.
Marine black shale deposits are common especially during the Dienerian substage of the Induan. These point to low oxygenation in the ocean.[12]
Induan life
The Induan age followed the mass extinction event at the end of the Permian period. Both global biodiversity and community-level (alpha) diversity remained low through much of this stage of the Triassic.[13]
Much of the supercontinent Pangea remained almost lifeless, deserted, hot, and dry. In higher latitudes, the flora during the Griesbachian was gymnosperm dominated but became lycopod dominated (e.g. Pleuromeia) in the Dienerian.[14] This change reflects a shift in global climate from cool and dry in the Griesbachian to hot and humid in the Dienerian and points to an extinction event during the Induan, just ca. 500'000 years after the end-Permian mass extinction event.[15] It led to the extinction of the Permian Glossopteris flora.
The lystrosaurids (below) and the proterosuchids (below) were the only groups of land animals to dominate during the Induan stage. Other animals, such as the ammonoids, insects, and the tetrapods (cynodonts, amphibians, reptiles, etc.) remained rare and terrestrial ecosystems did not recover for some 30 million years.[13] Both the seas and much of the freshwater during the Induan were anoxic, predominantly during the Dienerian subage.[12] Microbial reefs were common, possibly due to lack of competition with metazoan reef builders as a result of the extinction.[16]

Ray-finned fishes largely remained unaffected by the Permian-Triassic extinction event.[17][18] Many genera show a cosmopolitan (worldwide) distribution during the Induan and Olenekian (e.g. Australosomus, Birgeria, Parasemionotidae, Pteronisculus, Ptycholepidae, Saurichthys). This is well exemplified in the Griesbachian aged fish assemblages of the Wordie Creek Formation (East Greenland), the Dienerian aged assemblages of the Sakamena Formation (Madagascar), Candelaria Formation (Nevada, United States), and Mikin Formation (Himachal Pradesh, India), and the Smithian (Olenekian) aged assemblages of the Vikinghøgda Formation (Spitsbergen, Norway), Thaynes Formation (western United States), and Helongshan Formation (Anhui, China).
Induan Chondrichthyans include hybodonts, neoselachians and a few surviving lineages of eugeneodontid holocephalians, a mainly Palaeozoic group. Cartilaginous fishes were seemingly rare during the Induan.
Crocodile-shaped, marine temnospondyl amphibians (e.g. Aphaneramma, Wantzosaurus) were geographically widespread during the Induan and Olenekian ages. Their fossils are found in Greenland, Spitsbergen, Pakistan and Madagascar.[19]

The bivalve Claraia was widespread and common in the Panthalassa and Tethys oceans. The geologically oldest oysters (Liostrea) are known from the Induan. They grew on the shells of living ammonoids.[20]
- Conodonts
- Saurichthyids
- Birgeriids
- Ptycholepidae
- Parasemionotids
- Trematosaurids
- Lystrosaurids
- Proterosuchids
†Ammonoids
| †Ammonoidea of the Induan | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
|
worldwide | An gyronitid ammonoid |
 Ophiceras himalayanum | |
|
worldwide | An gyronitid ammonoid | ||
| worldwide | An ophiceratid ammonoid | |||
|
worldwide | An otoceratid ammonoid | ||
†Conodonts
| †Conodonta of the Induan | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| worldwide | index fossil for the base of the Triassic |
| ||

Cartilaginous fishes
| Chondrichthyes of the Induan | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Carboniferous to Early Triassic | United States, Canada | A eugeneodontid holocephalian (caseodontid) |
 Parahelicampodus | |
| Carboniferous to Early Triassic | United States, Canada, Greenland | A eugeneodontid holocephalian (caseodontid) | ||
| Early Triassic to Early Cretaceous | South Africa, Greenland, Svalbard | A hybodontiform elasmobranch | ||
| Carboniferous to Early Triassic | Canada | A chondrichthyan with uncertain affinities, known primarily from their feather-like denticles. | ||
|
Early Triassic | Greenland | A eugeneodontid holocephalian (edestid) | |



Ray-finned fishes
| Actinopterygii of the Induan | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | Nevada, United States | A non-neopterygian |
| |
| Early Triassic | Greenland, Madagascar | A non-neopterygian | ||
| Triassic | Greenland, Madagascar | A non-neopterygian | ||
| Lopingian to Middle Triassic | Greenland, Madagascar, Canada | A non-neopterygian | ||
| Early Triassic | Greenland, Madagascar | A non-neopterygian | ||
| Early Triassic | Nevada, United States | A neopterygian | ||
| Early Triassic | Greenland, Madagascar | A platysiagid | ||
| Early Triassic to Middle Triassic | Greenland, Madagascar, United States | A non-neopterygian | ||
| Triassic | Greenland, Madagascar | A non-neopterygian | ||
| Early Triassic | Madagascar | A neopterygian | ||


Coelacanths
| Actinistia of the Induan | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | Canada | A coelacanth. | ||
| Early Triassic | Greenland | A coelacanth | ||
| Early Triassic | Madagascar | A small coelacanth | ||
| Carboniferous to Early Triassic | Madagascar | A coelacanth | ||
| Early Triassic | Greenland | A coelacanth | ||
| Early Triassic | Greenland, Madagascar, Canada | A coelacanth | ||
Lungfishes
| Dipnoi of the Induan | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | Madagascar |
| ||
| Early Triassic | Angola and Madagascar | |||
| Early Triassic | Madagascar | |||
| Early Triassic | Australia | |||

†Temnospondyls
| Temnospondyli of the Induan | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | Pakistan; Madagascar | A trematosaurid stereospondyl amphibian |
| |
| Early Triassic | Australia | A brachyopid stereospondyl amphibian. | ||
| Beaufort Group, Karoo Basin, South Africa | A genus of stereospondyl amphibian in the family Rhinesuchidae. | |||
| Early Triassic | Knocklofty Formation, Australia | A lydekkerinid stereospondyl amphibian. | ||
| Early Triassic | Lower Fremouw Formation, Antarctica | A lydekkerinid stereospondyl amphibian. | ||
| Early Triassic | Middle Sakamena Formation, Madagascar | A lydekkerinid stereospondyl amphibian. | ||
| Induan to Smithian | Madagascar | A capitosaurian amphibian closely related to Watsonisuchus. | ||
| Early Triassic | Lystrosaurus Assemblage Zone, Katberg Formation, South Africa | A lydekkerinid stereospondyl amphibian. | ||
| Early Triassic | Queensland, Australia | A brachyopomorph stereospondyl amphibian. A possible synonym of the closely related genus Bothriceps. | ||
| Arcadia Formation, Australia | A basal stereospondyl amphibian in the family Lapillopsidae. | |||
| Early Triassic | South Africa; Australia. | A lydekkerinid stereospondyl amphibian. | ||
| West Bengal, India | A basal stereospondyl amphibian in the family Lapillopsidae. | |||
| Early Triassic | Lystrosaurus Assemblage Zone, Karoo Basin, South Africa | A genus of dissorophoid temnospondyl in the family Micropholidae. It was previously included in Amphibamidae along with other micropholids. However, a number of analyses recover Micropholidae as a distinct clade of derived non-olsoniform dissorophoids. | ||
| Australia | A stereospondyl amphibian in the family Rhytidosteidae. | |||
| Australia | A stereospondyl amphibian in the family Rhytidosteidae. | |||
| Early Induan to Early Olenekian | Knocklofty Formation, Tasmania | A basal stereospondyl amphibian in the family Lapillopsidae. | ||
| Early Triassic | Astrakhan Oblast, Russia | A rhytidosteid stereospondyl amphibian. | ||
| Early Triassic | Rio Grande do Sul, Brazil | A stereospondyl amphibian in the family Rhytidosteidae. | ||
| South Africa | A dvinosaurian amphibian within the family Tupilakosauridae. | |||
| Early Triassic | Greenland; Madagascar | A trematosaurid stereospondyl amphibian | ||
| Early Triassic | Australia; South Africa | A capitosaurian amphibian | ||
| Early Triassic | Queensland, Australia | A brachyopid stereospondyl amphibian. | ||






†Chroniosuchians
| Chroniosuchia of the Induan | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | Russia | A bystrowianid reptiliomorph |
| |
| Lopingian to Early Triassic | Russia, China | A reptiliomorph | ||


Lissamphibia
| Lissamphibians of the Induan | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Early Triassic | Madagascar | Triadobatrachus is an extinct genus of salientian frog-like amphibians, including only one known species, Triadobatrachus massinoti. It is the oldest member of the frog lineage known, and an excellent example of a transitional fossil. |
| |

†Procolophonomorphs
| Procolophonomorpha of the Induan | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Lopingian to Early Triassic | Madagascar | An owenettid parareptile. |
| |
| South Africa | A procolophonid parareptile, it was similar to owenettids due to its very basal position. | |||
| Early Triassic | China | A species of procolophonid parareptile, currently classified under the procolophonin genus Eumetabolodon though it is more basal and closely related to Theledectes. | ||
| Lopingian to Early Triassic | South Africa | An owenettid parareptile. | ||
| Vokhma Formation. Nizhnii Novgorod, Russia | A basal procolophonid parareptile. | |||
| Late Permian to Late Triassic | South Africa | A genus of lizard-shaped parareptile. | ||
| South Africa | A basal procolophonid parareptile. | |||
| Early Induan | South Africa | An owenettid parareptile. | ||
| South Africa | A basal procolophonid parareptile. | |||


Diapsids
| Diapsida of the Induan | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Late Permian to Early Triassic | Madagascar | An aquatic tangasaurid younginiform reptile. |
| |
| Early Triassic | Coast Province, southeastern Kenya. | A relatively small, lightly built, general lizard-like reptile, most likely a tangasaurid younginiform but without distinct aquatic adaptations. | ||
| Late Permian to Early Triassic | Katberg Formation, South Africa | An an extinct genus of lizard-like lepidosauromorph reptile. | ||

Archosauromorphs
| Archosauromorphs of the Induan | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| Arcadia Formation, Queensland, Australia. | A small basal archosauromorph, probably closely related to Prolacerta. |
| ||
| early Induan | Katberg Formation, Beaufort Group, South Africa | A basal rhynchosaur, the oldest known to date. | ||
| South Africa, Fremouw Formation, Antarctica | A small, lizard-like, basal archosauromorph, originally thought to be a lepidosauromorph. | |||
| South Africa | The largest land reptile during the Early Triassic period, equivalent in size to today's Komodo dragons. It looked somewhat similar to a primitive crocodile, and shared many of their modern features like long jaws, powerful neck muscles, short legs and a lengthy tail, while retaining several of its own unique features such as its long legs, and hooked shaped mouth. | |||
| Induan to Olenekian | Knocklofty Formation, West Hobart, Tasmania | Once believed to be a proterosuchid, this taxon is now believed to have been intermediate between advanced non-archosauriform archosauromorphs such as Prolacerta, and basal archosauriforms such as Proterosuchus. This genus is also notable being one of the most complete Australian Triassic reptiles known. | ||
| Induan to Olenekian | Southern Urals, Russia | Originally classified as a rauisuchid, Tsylmosuchus has more recently been interpreted as an indeterminate archosauriform. Tsylmosuchus occurred throughout the Olenekian age. Some of the strata from which Tsylmosuchus has been found are Induan in age, making it one of the earliest archosaurs. | ||
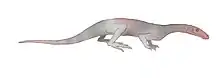

Therapsids
| Therapsids of the Induan | ||||
|---|---|---|---|---|
| Taxa | Presence | Location | Description | Images |
| A cynodont. |
| |||
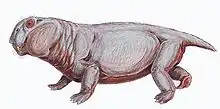
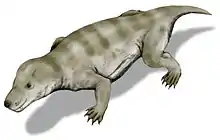
| Late Permian to Early Triassic | Antarctica, Russia, India and South Africa | A dicynodont. It was the most common group of terrestrial vertebrates during the Early Triassic, for a while 95% of land vertebrates were Lystrosaurus | ||
| Late Permian to Early Triassic | South Africa | A therocephalian. | ||
| A therocephalian. | ||||
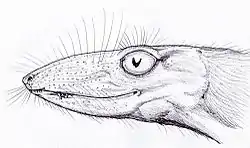
| Early Triassic | South Africa, Antarctica | A cat-sized cynodont. Many scientists suggest that the pits on the skull indicate that Thrinaxodon had whiskers and, therefore, probably had a covering of fur. There are suggestions that it was warm-blooded. Even so, it still laid eggs. | ||


References
- Widmann, Philipp; Bucher, Hugo; Leu, Marc; Vennemann, Torsten; Bagherpour, Borhan; Schneebeli-Hermann, Elke; Goudemand, Nicolas; Schaltegger, Urs (2020). "Dynamics of the Largest Carbon Isotope Excursion During the Early Triassic Biotic Recovery". Frontiers in Earth Science. 8 (196): 1–16. doi:10.3389/feart.2020.00196.
- McElwain, J. C.; Punyasena, S. W. (2007). "Mass extinction events and the plant fossil record". Trends in Ecology & Evolution. 22 (10): 548–557. doi:10.1016/j.tree.2007.09.003. PMID 17919771.
- |note5-nudge-down=1 Retallack, G. J.; Veevers, J.; Morante, R. (1996). "Global coal gap between Permian–Triassic extinctions and middle Triassic recovery of peat forming plants". GSA Bulletin. 108 (2): 195–207. doi:10.1130/0016-7606(1996)108<0195:GCGBPT>2.3.CO;2. Retrieved 2007-09-29.
- Payne, J. L.; Lehrmann, D. J.; Wei, J.; Orchard, M. J.; Schrag, D. P.; Knoll, A. H. (2004). "Large Perturbations of the Carbon Cycle During Recovery from the End-Permian Extinction". Science. 305 (5683): 506–9. doi:10.1126/science.1097023. PMID 15273391.
- Hongfu, Yin; Kexin, Zhang; Jinnan, Tong; Zunyi, Yang; Shunbao, Wu (June 2001). "The Global Stratotype Section and Point (GSSP) of the Permian-Triassic Boundary" (PDF). Episodes. 24 (2): 102–114. doi:10.18814/epiiugs/2001/v24i2/004. Retrieved 8 December 2020.
- "Global Boundary Stratotype Section and Point". International Commission of Stratigraphy. Retrieved 23 December 2020.
- http://www.stratigraphy.org/index.php/ics-chart-timescale
- Tozer E. T. (1965): Lower Triassic stages and ammonoid zones of Arctic Canada: Paper of the Geological Survey of Canada 65:1–14.
- Kiparisova & Popov (1956)
- The Triassic Timescale, Spencer Lucas (2010), ISBN 9781862392960
- Yin Hongfu, Zhang Kexin, Tong Jinnan, Yang Zunyi und Wu Shunbao: '"The Global Stratotype Section and Point (GSSP)of the Permian-Triassic Boundary." Episodes, 24(2): 102-114, Beijing 2001 ISSN 0705-3797.
- Ware et al. (2015): High-resolution biochronology and diversity dynamics of the Early Triassic ammonoid recovery: the Dienerian faunas of the Northern Indian Margin. Palaeogeography, Palaeoclimatology, Palaeoecology 440:363-373 https://doi.org/10.1016/j.palaeo.2015.09.013
- Sahney, S.; Benton, M.J. (2008). "Recovery from the most profound mass extinction of all time" (PDF). Proceedings of the Royal Society B: Biological Sciences. 275 (1636): 759–65. doi:10.1098/rspb.2007.1370. PMC 2596898. PMID 18198148.
- Schneebeli-Hermann et al. (2015): Vegetation history across the Permian–Triassic boundary in Pakistan (Amb section, Salt Range). Gondwana Research 27:911-924 http://dx.doi.org/10.1016/j.gr.2013.11.007
- Hochuli et al. (2016): Severest crisis overlooked—Worst disruption of terrestrial environments postdates the Permian–Triassic mass extinction. Scientific Reports 6:28372 https://doi.org/10.1038/srep28372
- Foster et al. (2020): Suppressed competitive exclusion enabled the proliferation of Permian/Triassic boundary microbialites. The Depositional record 6. 1–13. https://doi.org/10.1002/dep2.97
- Romano et al. (2016): Permian–Triassic Osteichthyes (bony fishes): diversity dynamics and body size evolutionBiological Reviews 91:106-147 https://doi.org/10.1111/brv.12161
- Smithwick F.M., and Stubbs T.L. (2018): Phanerozoic survivors: Actinopterygian evolution through the Permo‐Triassic and Triassic‐Jurassic mass extinction events. Evolution 72:348-362. https://doi.org/10.1111/evo.13421<
- Scheyer et al. (2014): Early Triassic Marine Biotic Recovery: The Predators' Perspective. PLoS ONE https://doi.org/10.1371/journal.pone.0088987
- Hautmann et al. (2017): Geologically oldest oysters were epizoans on Early Triassic ammonoids. Journal of Molluscan Studies 83:253-260 https://doi.org/10.1093/mollus/eyx018
Sources
- Brack, P.; Rieber, H.; Nicora, A. & Mundil, R.; 2005: The Global boundary Stratotype Section and Point (GSSP) of the Ladinian Stage (Middle Triassic) at Bagolino (Southern Alps, Northern Italy) and its implications for the Triassic time scale, Episodes 28(4), pp. 233–244.
- Gradstein, F. M.; Ogg, J. G. & Smith, A. G.; 2004: A Geologic Time Scale 2004, Cambridge University Press.
- Kiparisova, Lubov Dmitrievna & Popov, Yurij Nikolaivitch; 1956: Расчленение нижнего отдела триасовой системы на ярусы (Subdivision of the lower series of the Triassic System into stages), Doklady Akademii Nauk SSSR 109(4), pp 842–845 (in Russian).
External links
- GeoWhen Database - Induan
- Lower Triassic timescale at the website of the subcommission for stratigraphic information of the ICS
- Lower Triassic timescale at the website of Norges Network of offshore records of geology and stratigraphy.
