Alectrosaurus
Alectrosaurus (/əˌlɛktroʊˈsɔːrəs/; meaning "alone lizard") is a genus of tyrannosauroid theropod dinosaur that lived in Asia during the Late Cretaceous period, about some 96 million years ago in what is now the Iren Dabasu Formation.
| Alectrosaurus | |
|---|---|
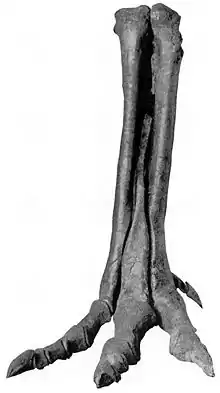 | |
| Right pes of A. olseni, specimen AMNH 6554, American Museum of Natural History | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | †Pantyrannosauria |
| Clade: | †Eutyrannosauria |
| Genus: | †Alectrosaurus Gilmore, 1933 |
| Type species | |
| †Alectrosaurus olseni Gilmore, 1933 | |
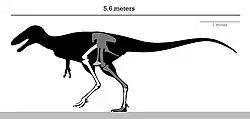
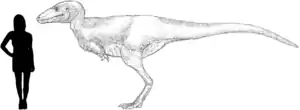
It was a medium-sized, moderately-built, ground-dwelling, bipedal carnivore, estimated at 5 to 6 m (16 to 20 ft) with a body shape similar to its much larger advanced relative, Tyrannosaurus. Alectrosaurus was a very fast running tyrannosauroid as indicated by the elongated hindlimbs that likely filled the niche of a pursuit predator, a trait that seems to be lost by the advanced and robust tyrannosaurids.
Discovery and naming

In 1923, the Third Asiatic Expedition of the American Museum of Natural History, led by chief paleontologist Walter W. Granger was hunting for dinosaur fossils in Mongolia. On April 25, assistant paleontologist George Olsen excavated and recovered the holotype AMNH 6554, a nearly complete right hindlimb. This included a virtually complete right hindlimb with some elements from the left pes and two manual unguals. On May 4, Olsen discovered another specimen approximately 30 m (98.4 ft) away from his first find, catalogued as AMNH 6368. This specimen included a right humerus, two incomplete manual digits, four fragmentary caudal vertebrae, and other two or three unspecified elements that were discarded due to badly preservation. These discoveries were made at the Iren Dabasu Formation in what is now the Inner Mongolia Autonomous Region (Nei Mongol Zizhiqu) of China.[1]
Both genus and species were formally described and named by the American paleontologist Charles Gilmore in 1933. The generic name, Alectrosaurus, can be translated as "alone lizard" or "mateless lizard", derived from the Greek words ἄλεκτρος (meaning alone or unmarried) and σαῦρος (meaning lizard). The specific name, olseni, is in honor of George Olsen, who discovered the first specimens.[1]
Since then, more material has been referred to Alectrosaurus. Possible findings of two additional specimens at the Bayan Shireh Formation were reported and described by the Mongolian paleontologist Altangerel Perle in 1977. The specimen IGM 100/50 consists of a partial maxilla, scapulocoracoid and manual ungual, and specimen IGM 100/51 consists of a fragmentary skull with lower jaws and other elements, incomplete ilium, and metatarsals of the right foot. These fossils were found in Outer Mongolia.[2][3] Iren Dabasu and Bayan Shireh dinosaur faunas are very similar, so it is not surprising that a species of Alectrosaurus would be found there. Furthermore, several partial skeletons found in both Inner and Outer Mongolia might belong to Alectrosaurus.[4] Alexander Averianov and Hans-Dieter Sues 2012 have estimated that the Iren Dabasu Formation is Santonian in age, correlating the Upper Bayan Shireh Formation.[5] Van Itterbeeck et al. 2005 suggested that the Iren Dabasu Formation is probably Campanian-Maastrichtian in age and possibly correlated with the Nemegt Formation.[6]
Near the holotype, the specimen AMNH 6556 was found in the same strata but at different points also in 1923. It consists of premaxillary and lateral teeth, incomplete left lacrimal, maxillary process of the left jugal, partial right quadratojugal, jugal process of the right ectopterygoid and the quadrate ramus to the right pterygoid. Although the specimen seems to represent a smaller individual. Its affinity to Alectrosaurus is somewhat unresolved since the specimen lacks hindlimb material, making direct comparisons with Alectrosaurus quite complicated.[7][8]
Description
The lectotype AMNH 6554 is fragmentary, consisting of a nearly complete right hindlimb only lacking the distal tarsal elements; left metatarsals II, III and IV, and a fragmentary distal foot of a pubis, however it is unknown which pubis represents.[1][3]
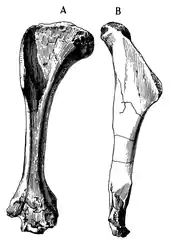
It was a medium-sized tyrannosauroid, reaching a length between 5 to 6 m (16 to 20 ft), and a weight ranging from 454 to 907 kg (1,001 to 2,000 lb).[9][10] Overall, the hindlimbs were rather gracile, in contrast to the robust tyrannosaurids. The length of its tibia (shinbone) and femur (thighbone) are very close, in contrast to the majority of other tyrannosauroids, where the tibia is longer. The femur measures 72.7 cm (727 mm) and the tibia 73 cm (730 mm). The metetarsals are also closer in size to the tibia than in most other tyrannosauroids, where they are usually longer; the third is the largest, measuring 48 cm (480 mm) long.[1][11] The astragalus and calcaneum are nicely preserved, although the astragalus seems to be slightly damaged. They are strongly attached, but not fused.[1]
Distinguishing anatomical features
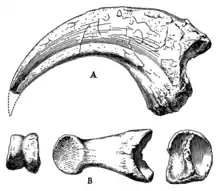
Following the original description of Alectrosaurus, it can be distinguished by the following traits: long slender-limbed type of tyrannosauroid; humerus long and slender; ungual and phalanx of digit I robust, laterally compressed and strongly curved; femur and tibia subequal in length; length of astragalus onefourth the combined length of astragalus and tibia.[1]
According to Carr 2005, Alectrosaurus can be distinguished based on unique traits present in the hindlimbs, such as the spike-like process extending from the caudodorsal surface of the medial condyle of the femur, the presence of an abrupt expansion in length of the anterior margin of the joint surface for the tibia on the fibula, tendon pit adjacent to the ventrolateral buttress of the astragalus undercutting the medial surface of the buttress, the dorsal margin of the proximal surface of pedal phalanx II-2 is pointed, reduced pedal digit III, the lateral condyle of pedal phalanx III-1 is significantly deeper than the medial condyle, when in distal view, stocky pedal phalanx IV-2, when examined in proximal view, the dorsal half of the joint surface for metatarsal IV on metatarsal III is dilated anteriorly, and many others.[12]
Classification
In 1933, Charles Gilmore examined the available material and concluded that AMNH 6554 and AMNH 6368 were syntypes belonging to the same genus. He based this on his observation that the manual unguals from both specimens were morphologically similar. Observing similarities with the hindlimbs of specimen AMNH 5664 Gorgosaurus sternbergi, he classified this new genus as a "Deinodont", a term that is now considered equivalent to tyrannosaurid.[1] Due to its fragmentary nature, there is presently very little confidence in restoring its relationships with other tyrannosauroids and many recent cladistic analyses have omitted it altogether. One study recovered Alectrosaurus at no less than eight equally parsimonious positions in a tyrannosauroid cladogram.[13] Some paleontologists have equivocally considered Alectrosaurus olseni to be a species of Albertosaurus.[14]
Alectrosaurus was originally characterized as a long-armed theropod, but Perle 1977 and Mader & Bradley 1989 observed that the forelimbs of the specimen AMNH 6368 did not belong to the genus, as they do not share characteristics with Tyrannosauroidea, and assigned them to the Therizinosauria incertae sedis.[2][3] The remaining material, AMNH 6554 represents the hind limb with characteristics of a true tyrannosauroid, and were assigned as the lectotype for Alectrosaurus olseni.[3] Additionally, four small caudal vertebrae were associated with the specimen AMNH 6368, the vertebrae were not included in the original description. Nevertheless, in 1984 they were catalogued as AMNH 21784. Mader and Bradley described these vertebrae, and were provisionally identified as caudal vertebrae of a small theropod dinosaur that is not phylogenetically referable to either the Tyrannosauroidea or Therizinosauridae as they show resemblance to the caudal vertebrae of Deinonychus and Plateosaurus.[3]
The Bayan Shireh material may or may not belong to this genus, and needs further study. One cladistic analysis showed that the two sets of specimens group together exclusive of any other taxa, so they are probably at least closely related, if not the same species.[15]
In 2018, Rafael Delcourt and Orlando Nelson Grillo published an extensive phylogenetic analysis of the Tyrannosauroidea, classifying Gondwanan and Laurasian tyrannosaurs. Below is the cladogram showing the phylogenetic position of Alectrosaurus according to their analysis:[16]
| Tyrannosauroidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Palaeobiology

The hindlimb of the specimen AMNH 6554 is notable for the particular elongated digits and metatarsals, differing from other tyrannosauroids. These traits are found in terrestrial runner birds, suggesting that Alectrosaurus was suited as a fast-running tyrannosauroid dinosaur with well developed hindlimbs; probably a pursuit predator.[7] This interpretation is consistent with the results obtained in the limb proportion analysis performed by Scott Persons IV and Currie in 2016. By comparing the limbs of numerous theropods they noted that most tyrannosauroids were highly cursorial and leggy animals, with the exception of giant and stocky-legged forms such as Tarbosaurus or Tyrannosaurus. Alectrosaurus was recovered with a relatively high CLP (Cursorial-limb-proportion) score at 16.5, higher than most carnosaurs.[11] In 2001, a study conducted by Bruce Rothschild and colleagues, examined 23 foot bones referred to Alectrosaurus for signs of stress fractures, but none were found.[17]
Paleoecology

Alectrosaurus was first recovered from the Iren Dabasu Formation, which has been dated to the Cenomanian stage, about 95.8 ± 6.2 million years ago.[18] During the Late Cretaceous, there was a large floodplain with braided fluvial environments in the formation. The floodplain environments had extensive vegetation, evidenced in the palaeosol development and the numerous herbivorous dinosaurs that were found in both the river channel and the floodplain sediments.[6] Contemporaneous paleofauna from this formation included another theropods such as Archaeornithomimus, Caenagnathasia, Erliansaurus, Gigantoraptor and Neimongosaurus; the sauropod Sonidosaurus and the two hadrosauroids Bactrosaurus and Gilmoreosaurus. Alectrosaurus likely preyed on these two.[19][20] Additional to this, a potential discovery was made on the Bayan Shireh Formation at the Bayshi Tsav locality.[2] The Bayan Shireh Formation is estimated to be Late Cretaceous in age, during the Cenomanian-Santonian stages, about 95.9 ± 6.0 million to 89.6 ± 4.0 million years ago, similar to Iren Dabasu.[21][22] Here, Alectrosaurus lived alongside diverse theropods such as Achillobator, Garudimimus or Segnosaurus. Ankylosaurs were represented by Talarurus and Tsagantegia, other herbivorous dinosaurs include Graciliceratops, Gobihadros and Erketu.[23][24][25]
Alexander Averianov and Hans-Dieter Sues in 2012 estimated that the Upper Bayan Shireh correlated the Iren Dabasu Formation and both had similar environments. This correlation seems to be supported by a large number of ostracods reported in these formations.[5] Further support can be evidenced on the similarities between the dinosaur taxa. As previously mentioned, Alectrosaurus appears to occur in both Iren Dabasu and Bayan Shireh Formations, although this referral to the genus has not been fully confirmed, the tentative specimens show striking resemblance to the Iren Dabasu material. In 2015, a giant caenagnathid was reported from the Bayan Shireh Formation, preserving a partial lower rostrum (beak) that is extremely similar to that of Gigantotaptor. If confirmed by future studies, this represents the first documented occurrence of a taxon in both formations.[2][26]
See also
| Wikimedia Commons has media related to Alectrosaurus. |
References
- Gilmore, C. W. (1933). "On the dinosaurian fauna of the Iren Dabasu Formation". Bulletin of the American Museum of Natural History. 67 (2): 23–78. hdl:2246/355.
- Perle, A. (1977). "O pervoy nakhodke Alektrozavra (Tyrannosauridae, Theropoda) iz pozdnego Mela Mongolii" [On the first discovery of Alectrosaurus (Tyrannosauridae, Theropoda) in the Late Cretaceous of Mongolia]. Shinzhlekh Ukhaany Akademi Geologiin Khureelen (in Russian). 3 (3): 104–113.
- Mader, B. J.; Bradley, R. L. (1989). "A redescription and revised diagnosis of the syntypes of the Mongolian tyrannosaur Alectrosaurus olseni". Journal of Vertebrate Paleontology. 9 (1): 41–55. doi:10.1080/02724634.1989.10011737.
- Benton, M. J.; Shishkin, M. A.; Unwin, D. M.; Kurochkin, E. N. (2003). The Age of Dinosaurs in Russia and Mongolia. Cambridge University Press. pp. 434–455. ISBN 0-521-54582-X.
- Averianov, A.; Sues, H. (2012). "Correlation of Late Cretaceous continental vertebrate assemblages in Middle and Central Asia" (PDF). Journal of Stratigraphy. 36 (2): 462–485.
- Van Itterbeeck, J.; Horne, D. J.; Bultynck, P.; Vandenberghe, N. (2005). "Stratigraphy and palaeoenvironment of the dinosaur-bearing Upper Cretaceous Iren Dabasu Formation, Inner Mongolia, People's Republic of China". Cretaceous Research. 26 (4): 699–725. doi:10.1016/j.cretres.2005.03.004.
- Carr, T. D.; Williamson, T. E. (2005). "A reappraisal of tyrannosauroids from Iren Dabasu, Inner Mongolia, People's Republic of China". Journal of Vertebrate Paleontology. 25 (3).
- Carr, T. D.; Varricchio, D. J.; Sedlmayr, J. C.; Roberts, E. M.; Moore, J. R. (2017). "A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system". Scientific Reports. 7 (44942): 44942. Bibcode:2017NatSR...744942C. doi:10.1038/srep44942. PMC 5372470. PMID 28358353.
- Holtz, T. R.; Rey, L. V. (2007). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Random House. Genus List for Holtz 2012 Weight Information
- Brett-Surman, M. K.; Holtz, T. R.; Farlow, J. O. (2012). The Complete Dinosaur. Indiana University Press. p. 360. ISBN 978-0-2533-5701-4.
- Scott Persons IV, W.; Currie, P. J. (2016). "An approach to scoring cursorial limb proportions in carnivorous dinosaurs and an attempt to account for allometry". Scientific Reports. 6 (19828): 19828. Bibcode:2016NatSR...619828P. doi:10.1038/srep19828. PMC 4728391. PMID 26813782.
- Carr, T. D. (2005). Phylogeny of Tyrannosauroidea (Dinosauria: Coelurosauria) with Special Reference to North American Forms. University of Toronto. p. 1170. ISBN 0-494-02932-3. OCLC 75087116.
- Weishampel, D. B.; Dodson, P.; Osmolska, H. (2007). The Dinosauria, Second Edition. University of California Press. pp. 111–136. ISBN 978-0-520-24209-8.
- Dodson, P.; Britt, B.; Carpenter, K.; Forster, C. A.; Gillete, D. D.; Norell, M. A.; Olshevsky, G.; Parrish, J. M.; Weishampel, D. B. (1994). "Albertosaurus". The Age of Dinosaurs. Publications International, Ltd. pp. 106–107. ISBN 0-7853-0443-6.
- Holtz, T. R. (2001). "The phylogeny and taxonomy of the Tyrannosauridae". Mesozoic Vertebrate Life. Indiana University Press. pp. 64–83. ISBN 0-253-33907-3.
- Delcourt, R.; Grillo, O. N. (2018). "Tyrannosauroids from the Southern Hemisphere: Implications for biogeography, evolution, and taxonomy". Palaeogeography, Palaeoclimatology, Palaeoecology. 511: 379–387. Bibcode:2018PPP...511..379D. doi:10.1016/j.palaeo.2018.09.003.
- Rothschild, B.; Tanke, D.; Ford, T. (2001). "Theropod stress fractures and tendon avulsions as a clue to activity". Mesozoic Vertebrate Life. Indiana University Press. pp. 331–336.
- Guo, Z. X.; Shi, Y. P.; Yang, Y. T.; Jiang, S. Q.; Li, L. B.; Zhao, Z. G. (2018). "Inversion of the Erlian Basin (NE China) in the early Late Cretaceous: Implications for the collision of the Okhotomorsk Block with East Asia" (PDF). Journal of Asian Earth Sciences. 154: 49–66. doi:10.1016/j.jseaes.2017.12.007.
- Xing, H.; He, Y.; Li, L.; Xi, D. (2012). "A review on the study of the stratigraphy, sedimentology, and paleontology of the Iren Dabasu Formation, Inner Mongolia". In Wei, D. (ed.). Proceedings of the Thirteenth Annual Meeting of the Chinese Society of Vertebrate Paleontology (in Chinese). Beijing: China Ocean Press. pp. 1–44.
- Xi, Y.; Xiao-Li, W.; Sullivan, C.; Shuo, W.; Stidham, T.; Xing, X. (2015). "Caenagnathasia sp. (Theropoda: Oviraptorosauria) from the Iren Dabasu Formation (Upper Cretaceous: Campanian) of Erenhot, Nei Mongol, China" (PDF). Vertebrata PalAsiatica. 53 (4): 291 298.
- Hicks, J. F.; Brinkman, D. L.; Nichols, D. J.; Watabe, M. (1999). "Paleomagnetic and palynologic analyses of Albian to Santonian strata at Bayn Shireh, Burkhant, and Khuren Dukh, eastern Gobi Desert, Mongolia". Cretaceous Research. 20 (6): 829–850. doi:10.1006/cres.1999.0188.
- Kurumada, Y.; Aoki, S.; Aoki, K.; Kato, D.; Saneyoshi, M.; Tsogtbaatar, K.; Windley, B. F.; Ishigaki, S. (2020). "Calcite U–Pb age of the Cretaceous vertebrate‐bearing Bayn Shire Formation in the Eastern Gobi Desert of Mongolia: usefulness of caliche for age determination". Terra Nova. doi:10.1111/ter.12456.
- Weishampel, D. B.; Barrett, P. M.; Coria, R. A.; Loeuff, J. L.; Xing, X.; Xijin, Z.; Sahni, A.; Gomani, E. M. P. (2004). "Dinosaur Distribution". In Weishampel, D. B.; Dodson, P.; Osmolska, H. (eds.). The Dinosauria (2nd ed.). University of California Press. pp. 596–598. ISBN 0520242092.
- Tsogtbaatar, K.; Weishampel, D. B.; Evans, D. C.; Watabe, M. (2019). "A new hadrosauroid (Dinosauria: Ornithopoda) from the Late Cretaceous Baynshire Formation of the Gobi Desert (Mongolia)". PLOS ONE. 14 (4): e0208480. Bibcode:2019PLoSO..1408480T. doi:10.1371/journal.pone.0208480. PMC 6469754. PMID 30995236.
- Park, J. (2020). "Additional skulls of Talarurus plicatospineus (Dinosauria: Ankylosauridae) and implications for paleobiogeography and paleoecology of armored dinosaurs". Cretaceous Research. 108: e104340. doi:10.1016/j.cretres.2019.104340.
- Tsuihiji, T.; Watabe, M.; Barsbold, R.; Tsogtbaatar, K. (2015). "A gigantic caenagnathid oviraptorosaurian (Dinosauria: Theropoda) from the Upper Cretaceous of the Gobi Desert, Mongolia". Cretaceous Research. 56: 60–65. doi:10.1016/j.cretres.2015.03.007.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||