Bee
Bees are flying insects closely related to wasps and ants, known for their role in pollination and, in the case of the best-known bee species, the western honey bee, for producing honey. Bees are a monophyletic lineage within the superfamily Apoidea. They are presently considered a clade, called Anthophila. There are over 16,000 known species of bees in seven recognized biological families.[1][2] Some species – including honey bees, bumblebees, and stingless bees – live socially in colonies while some species – including mason bees, carpenter bees, leafcutter bees, and sweat bees – are solitary.
| Bees | |
|---|---|
.jpg.webp) | |
| The sugarbag bee, Tetragonula carbonaria | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hymenoptera |
| (unranked): | Unicalcarida |
| Suborder: | Apocrita |
| Superfamily: | Apoidea |
| Clade: | Anthophila |
| Families | |
| Synonyms | |
|
Apiformes (from Latin 'apis') | |
Bees are found on every continent except for Antarctica, in every habitat on the planet that contains insect-pollinated flowering plants. The most common bees in the Northern Hemisphere are the Halictidae, or sweat bees, but they are small and often mistaken for wasps or flies. Bees range in size from tiny stingless bee species, whose workers are less than 2 millimetres (0.08 in) long, to Megachile pluto, the largest species of leafcutter bee, whose females can attain a length of 39 millimetres (1.54 in).
Bees feed on nectar and pollen, the former primarily as an energy source and the latter primarily for protein and other nutrients. Most pollen is used as food for their larvae. Vertebrate predators of bees include birds such as bee-eaters; insect predators include beewolves and dragonflies.
Bee pollination is important both ecologically and commercially, and the decline in wild bees has increased the value of pollination by commercially managed hives of honey bees. The analysis of 353 wild bee and hoverfly species across Britain from 1980 to 2013 found the insects have been lost from a quarter of the places they inhabited in 1980.[3]
Human beekeeping or apiculture has been practised for millennia, since at least the times of Ancient Egypt and Ancient Greece. Bees have appeared in mythology and folklore, through all phases of art and literature from ancient times to the present day, although primarily focused in the Northern Hemisphere where beekeeping is far more common.
Evolution

The ancestors of bees were wasps in the family Crabronidae, which were predators of other insects. The switch from insect prey to pollen may have resulted from the consumption of prey insects which were flower visitors and were partially covered with pollen when they were fed to the wasp larvae. This same evolutionary scenario may have occurred within the vespoid wasps, where the pollen wasps evolved from predatory ancestors. Until recently, the oldest non-compression bee fossil had been found in New Jersey amber, Cretotrigona prisca of Cretaceous age, a corbiculate bee.[4] A bee fossil from the early Cretaceous (~100 mya), Melittosphex burmensis, is considered "an extinct lineage of pollen-collecting Apoidea sister to the modern bees".[5] Derived features of its morphology (apomorphies) place it clearly within the bees, but it retains two unmodified ancestral traits (plesiomorphies) of the legs (two mid-tibial spurs, and a slender hind basitarsus), showing its transitional status.[5] By the Eocene (~45 mya) there was already considerable diversity among eusocial bee lineages.[6][lower-alpha 1]
The highly eusocial corbiculate Apidae appeared roughly 87 Mya, and the Allodapini (within the Apidae) around 53 Mya.[9] The Colletidae appear as fossils only from the late Oligocene (~25 Mya) to early Miocene.[10] The Melittidae are known from Palaeomacropis eocenicus in the Early Eocene.[11] The Megachilidae are known from trace fossils (characteristic leaf cuttings) from the Middle Eocene.[12] The Andrenidae are known from the Eocene-Oligocene boundary, around 34 Mya, of the Florissant shale.[13] The Halictidae first appear in the Early Eocene[14] with species[15][16] found in amber. The Stenotritidae are known from fossil brood cells of Pleistocene age.[17]
Coevolution

The earliest animal-pollinated flowers were shallow, cup-shaped blooms pollinated by insects such as beetles, so the syndrome of insect pollination was well established before the first appearance of bees. The novelty is that bees are specialized as pollination agents, with behavioral and physical modifications that specifically enhance pollination, and are the most efficient pollinating insects. In a process of coevolution, flowers developed floral rewards[18] such as nectar and longer tubes, and bees developed longer tongues to extract the nectar.[19] Bees also developed structures known as scopal hairs and pollen baskets to collect and carry pollen. The location and type differ among and between groups of bees. Most species have scopal hairs on their hind legs or on the underside of their abdomens. Some species in the family Apidae have pollen baskets on their hind legs, while very few lack these and instead collect pollen in their crops.[2] The appearance of these structures drove the adaptive radiation of the angiosperms, and, in turn, bees themselves.[7] Bees coevolved not only with flowers but it is believed that some species coevolved with mites. Some provide tufts of hairs called acarinaria that appear to provide lodgings for mites; in return, it is believed that mites eat fungi that attack pollen, so the relationship in this case may be mutualistc.[20][21]
External
This phylogenetic tree is based on Debevic et al, 2012, which used molecular phylogeny to demonstrate that the bees (Anthophila) arose from deep within the Crabronidae, which is therefore paraphyletic. The placement of the Heterogynaidae is uncertain.[22] The small subfamily Mellininae was not included in this analysis.
| Apoidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||


Internal
This cladogram of the bee families is based on Hedtke et al., 2013, which places the former families Dasypodaidae and Meganomiidae as subfamilies inside the Melittidae.[23] English names, where available, are given in parentheses.
| Anthophila (bees) |
| |||||||||||||||||||||||||||||||||||||||

.jpg.webp)


Characteristics
_lapping_mouthparts%252C_showing_labium_and_maxillae..jpg.webp)
Bees differ from closely related groups such as wasps by having branched or plume-like setae (hairs), combs on the forelimbs for cleaning their antennae, small anatomical differences in limb structure, and the venation of the hind wings; and in females, by having the seventh dorsal abdominal plate divided into two half-plates.[24]
Bees have the following characteristics:
- A pair of large compound eyes which cover much of the surface of the head. Between and above these are three small simple eyes (ocelli) which provide information on light intensity.
- The antennae usually have 13 segments in males and 12 in females, and are geniculate, having an elbow joint part way along. They house large numbers of sense organs that can detect touch (mechanoreceptors), smell and taste; and small, hairlike mechanoreceptors that can detect air movement so as to "hear" sounds.
- The mouthparts are adapted for both chewing and sucking by having both a pair of mandibles and a long proboscis for sucking up nectar.[25]
- The thorax has three segments, each with a pair of robust legs, and a pair of membranous wings on the hind two segments. The front legs of corbiculate bees bear combs for cleaning the antennae, and in many species the hind legs bear pollen baskets, flattened sections with incurving hairs to secure the collected pollen. The wings are synchronised in flight, and the somewhat smaller hind wings connect to the forewings by a row of hooks along their margin which connect to a groove in the forewing.
- The abdomen has nine segments, the hindermost three being modified into the sting.[25]

The largest species of bee is thought to be Wallace's giant bee Megachile pluto, whose females can attain a length of 39 millimetres (1.54 in).[26] The smallest species may be dwarf stingless bees in the tribe Meliponini whose workers are less than 2 millimetres (0.08 in) in length.[27]
Sociality
Haplodiploid breeding system

According to inclusive fitness theory, organisms can gain fitness not just through increasing their own reproductive output, but also that of close relatives. In evolutionary terms, individuals should help relatives when Cost < Relatedness * Benefit. The requirements for eusociality are more easily fulfilled by haplodiploid species such as bees because of their unusual relatedness structure.[28]
In haplodiploid species, females develop from fertilized eggs and males from unfertilized eggs. Because a male is haploid (has only one copy of each gene), his daughters (which are diploid, with two copies of each gene) share 100% of his genes and 50% of their mother's. Therefore, they share 75% of their genes with each other. This mechanism of sex determination gives rise to what W. D. Hamilton termed "supersisters", more closely related to their sisters than they would be to their own offspring.[29] Workers often do not reproduce, but they can pass on more of their genes by helping to raise their sisters (as queens) than they would by having their own offspring (each of which would only have 50% of their genes), assuming they would produce similar numbers. This unusual situation has been proposed as an explanation of the multiple (at least 9) evolutions of eusociality within Hymenoptera.[30][31]
Haplodiploidy is neither necessary nor sufficient for eusociality. Some eusocial species such as termites are not haplodiploid. Conversely, all bees are haplodiploid but not all are eusocial, and among eusocial species many queens mate with multiple males, creating half-sisters that share only 25% of each-other's genes.[32] But, monogamy (queens mating singly) is the ancestral state for all eusocial species so far investigated, so it is likely that haplodiploidy contributed to the evolution of eusociality in bees.[30]
Eusociality


Bees may be solitary or may live in various types of communities. Eusociality appears to have originated from at least three independent origins in halictid bees.[33] The most advanced of these are species with eusocial colonies; these are characterised by cooperative brood care and a division of labour into reproductive and non-reproductive adults, plus overlapping generations.[34] This division of labour creates specialized groups within eusocial societies which are called castes. In some species, groups of cohabiting females may be sisters, and if there is a division of labour within the group, they are considered semisocial. The group is called eusocial if, in addition, the group consists of a mother (the queen) and her daughters (workers). When the castes are purely behavioural alternatives, with no morphological differentiation other than size, the system is considered primitively eusocial, as in many paper wasps; when the castes are morphologically discrete, the system is considered highly eusocial.[19]
True honey bees (genus Apis, of which seven species are currently recognized) are highly eusocial, and are among the best known insects. Their colonies are established by swarms, consisting of a queen and several hundred workers. There are 29 subspecies of one of these species, Apis mellifera, native to Europe, the Middle East, and Africa. Africanized bees are a hybrid strain of A. mellifera that escaped from experiments involving crossing European and African subspecies; they are extremely defensive.[35]
Stingless bees are also highly eusocial. They practise mass provisioning, with complex nest architecture and perennial colonies also established via swarming.[36]

Many bumblebees are eusocial, similar to the eusocial Vespidae such as hornets in that the queen initiates a nest on her own rather than by swarming. Bumblebee colonies typically have from 50 to 200 bees at peak population, which occurs in mid to late summer. Nest architecture is simple, limited by the size of the pre-existing nest cavity, and colonies rarely last more than a year.[37] In 2011, the International Union for Conservation of Nature set up the Bumblebee Specialist Group to review the threat status of all bumblebee species worldwide using the IUCN Red List criteria.[38]
There are many more species of primitively eusocial than highly eusocial bees, but they have been studied less often. Most are in the family Halictidae, or "sweat bees". Colonies are typically small, with a dozen or fewer workers, on average. Queens and workers differ only in size, if at all. Most species have a single season colony cycle, even in the tropics, and only mated females hibernate. A few species have long active seasons and attain colony sizes in the hundreds, such as Halictus hesperus.[39] Some species are eusocial in parts of their range and solitary in others,[40] or have a mix of eusocial and solitary nests in the same population.[41] The orchid bees (Apidae) include some primitively eusocial species with similar biology. Some allodapine bees (Apidae) form primitively eusocial colonies, with progressive provisioning: a larva's food is supplied gradually as it develops, as is the case in honey bees and some bumblebees.[42]
Solitary and communal bees

Most other bees, including familiar insects such as carpenter bees, leafcutter bees and mason bees are solitary in the sense that every female is fertile, and typically inhabits a nest she constructs herself. There is no division of labor so these nests lack queens and worker bees for these species. Solitary bees typically produce neither honey nor beeswax. Bees collect pollen to feed their young, and have the necessary adaptations to do this. However, certain wasp species such as pollen wasps have similar behaviours, and a few species of bee scavenge from carcases to feed their offspring.[24] Solitary bees are important pollinators; they gather pollen to provision their nests with food for their brood. Often it is mixed with nectar to form a paste-like consistency. Some solitary bees have advanced types of pollen-carrying structures on their bodies. Very few species of solitary bee are being cultured for commercial pollination. Most of these species belong to a distinct set of genera which are commonly known by their nesting behavior or preferences, namely: carpenter bees, sweat bees, mason bees, plasterer bees, squash bees, dwarf carpenter bees, leafcutter bees, alkali bees and digger bees.[43]

Most solitary bees nest in the ground in a variety of soil textures and conditions while others create nests in hollow reeds or twigs, holes in wood. The female typically creates a compartment (a "cell") with an egg and some provisions for the resulting larva, then seals it off. A nest may consist of numerous cells. When the nest is in wood, usually the last (those closer to the entrance) contain eggs that will become males. The adult does not provide care for the brood once the egg is laid, and usually dies after making one or more nests. The males typically emerge first and are ready for mating when the females emerge. Solitary bees are either stingless or very unlikely to sting (only in self-defense, if ever).[44][45]

While solitary, females each make individual nests.[46] Some species, such as the European mason bee Hoplitis anthocopoides,[47] and the Dawson's Burrowing bee, Amegilla dawsoni,[48] are gregarious, preferring to make nests near others of the same species, and giving the appearance of being social. Large groups of solitary bee nests are called aggregations, to distinguish them from colonies. In some species, multiple females share a common nest, but each makes and provisions her own cells independently. This type of group is called "communal" and is not uncommon. The primary advantage appears to be that a nest entrance is easier to defend from predators and parasites when multiple females use that same entrance regularly.[47]
Biology
Life cycle
The life cycle of a bee, be it a solitary or social species, involves the laying of an egg, the development through several moults of a legless larva, a pupation stage during which the insect undergoes complete metamorphosis, followed by the emergence of a winged adult. Most solitary bees and bumble bees in temperate climates overwinter as adults or pupae and emerge in spring when increasing numbers of flowering plants come into bloom. The males usually emerge first and search for females with which to mate. The sex of a bee is determined by whether or not the egg is fertilised; after mating, a female stores the sperm, and determines which sex is required at the time each individual egg is laid, fertilised eggs producing female offspring and unfertilised eggs, males. Tropical bees may have several generations in a year and no diapause stage.[49][50][51][52]
The egg is generally oblong, slightly curved and tapering at one end. Solitary bees, lay each egg in a separate cell with a supply of mixed pollen and nectar next to it. This may be rolled into a pellet or placed in a pile and is known as mass provisioning. Social bee species provision progressively, that is, they feed the larva regularly while it grows. The nest varies from a hole in the ground or in wood, in solitary bees, to a substantial structure with wax combs in bumblebees and honey bees.[53]
In most species, larvae are whitish grubs, roughly oval and bluntly-pointed at both ends. They have 15 segments and spiracles in each segment for breathing. They have no legs but move within the cell, helped by tubercles on their sides. They have short horns on the head, jaws for chewing food and an appendage on either side of the mouth tipped with a bristle. There is a gland under the mouth that secretes a viscous liquid which solidifies into the silk they use to produce a cocoon. The cocoon is semi-transparent and the pupa can be seen through it. Over the course of a few days, the larva undergoes metamorphosis into a winged adult. When ready to emerge, the adult splits its skin dorsally and climbs out of the exuviae and breaks out of the cell.[53]
 Nest of common carder bumblebee, wax canopy removed to show winged workers and pupae in irregularly placed wax cells
Nest of common carder bumblebee, wax canopy removed to show winged workers and pupae in irregularly placed wax cells Carpenter bee nests in a cedar wood beam (sawn open)
Carpenter bee nests in a cedar wood beam (sawn open) Honeybees on brood comb with eggs and larvae in cells
Honeybees on brood comb with eggs and larvae in cells
Flight

Antoine Magnan's 1934 book Le vol des insectes, says that he and André Sainte-Laguë had applied the equations of air resistance to insects and found that their flight could not be explained by fixed-wing calculations, but that "One shouldn't be surprised that the results of the calculations don't square with reality".[54] This has led to a common misconception that bees "violate aerodynamic theory". In fact it merely confirms that bees do not engage in fixed-wing flight, and that their flight is explained by other mechanics, such as those used by helicopters.[55] In 1996 it was shown that vortices created by many insects' wings helped to provide lift.[56] High-speed cinematography[57] and robotic mock-up of a bee wing[58] showed that lift was generated by "the unconventional combination of short, choppy wing strokes, a rapid rotation of the wing as it flops over and reverses direction, and a very fast wing-beat frequency". Wing-beat frequency normally increases as size decreases, but as the bee's wing beat covers such a small arc, it flaps approximately 230 times per second, faster than a fruitfly (200 times per second) which is 80 times smaller.[59]
Navigation, communication, and finding food
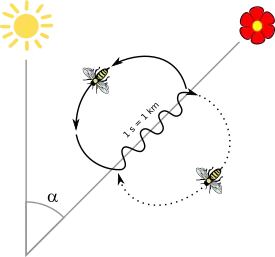
The ethologist Karl von Frisch studied navigation in the honey bee. He showed that honey bees communicate by the waggle dance, in which a worker indicates the location of a food source to other workers in the hive. He demonstrated that bees can recognize a desired compass direction in three different ways: by the sun, by the polarization pattern of the blue sky, and by the earth's magnetic field. He showed that the sun is the preferred or main compass; the other mechanisms are used under cloudy skies or inside a dark beehive.[60] Bees navigate using spatial memory with a "rich, map-like organization".[61]
Digestion
The gut of bees is relatively simple, but multiple metabolic strategies exist in the gut microbiota.[62] Pollinating bees consume nectar and pollen, which require different digestion strategies by somewhat specialized bacteria. While nectar is a liquid of mostly monosaccharide sugars and so easily absorbed, pollen contains complex polysaccharides: branching pectin and hemicellulose.[63] Approximately five groups of bacteria are involved in digestion. Three groups specialize in simple sugars (Snodgrassella and two groups of Lactobacillus), and two other groups in complex sugars (Gilliamella and Bifidobacterium). Digestion of pectin and hemicellulose is dominated by bacterial clades Gilliamella and Bifidobacterium respectively. Bacteria that cannot digest polysaccharides obtain enzymes from their neighbors, and bacteria that lack certain amino acids do the same, creating multiple ecological niches.[64]
Although most bee species are nectarivorous and palynivorous, some are not. Particularly unusual are vulture bees in the genus Trigona, which consume carrion and wasp brood, turning meat into a honey-like substance.[65]
Ecology
Floral relationships
Most bees are polylectic (generalist) meaning they collect pollen from a range of flowering plants, but some are oligoleges (specialists), in that they only gather pollen from one or a few species or genera of closely related plants.[66] Specialist pollinators also include bee species which gather floral oils instead of pollen, and male orchid bees, which gather aromatic compounds from orchids (one of the few cases where male bees are effective pollinators). Bees are able to sense the presence of desirable flowers through ultraviolet patterning on flowers, floral odors,[67] and even electromagnetic fields.[68] Once landed, a bee then uses nectar quality[67] and pollen taste[69] to determine whether to continue visiting similar flowers.
In rare cases, a plant species may only be effectively pollinated by a single bee species, and some plants are endangered at least in part because their pollinator is also threatened. But, there is a pronounced tendency for oligolectic bees to be associated with common, widespread plants visited by multiple pollinator species. For example, the creosote bush in the arid parts of the United States southwest is associated with some 40 oligoleges.[70]
As mimics and models


Many bees are aposematically coloured, typically orange and black, warning of their ability to defend themselves with a powerful sting. As such they are models for Batesian mimicry by non-stinging insects such as bee-flies, robber flies and hoverflies,[71] all of which gain a measure of protection by superficially looking and behaving like bees.[71]
Bees are themselves Müllerian mimics of other aposematic insects with the same colour scheme, including wasps, lycid and other beetles, and many butterflies and moths (Lepidoptera) which are themselves distasteful, often through acquiring bitter and poisonous chemicals from their plant food. All the Müllerian mimics, including bees, benefit from the reduced risk of predation that results from their easily recognised warning coloration.[72]
Bees are also mimicked by plants such as the bee orchid which imitates both the appearance and the scent of a female bee; male bees attempt to mate (pseudocopulation) with the furry lip of the flower, thus pollinating it.[73]
As brood parasites

Brood parasites occur in several bee families including the apid subfamily Nomadinae.[74] Females of these species lack pollen collecting structures (the scopa) and do not construct their own nests. They typically enter the nests of pollen collecting species, and lay their eggs in cells provisioned by the host bee. When the "cuckoo" bee larva hatches, it consumes the host larva's pollen ball, and often the host egg also.[75] In particular, the Arctic bee species, Bombus hyperboreus is an aggressive species that attacks and enslaves other bees of the same subgenus. However, unlike many other bee brood parasites, they have pollen baskets and often collect pollen.[76]
In Southern Africa, hives of African honeybees (A. mellifera scutellata) are being destroyed by parasitic workers of the Cape honeybee, A. m. capensis. These lay diploid eggs ("thelytoky"), escaping normal worker policing, leading to the colony's destruction; the parasites can then move to other hives.[77]
The cuckoo bees in the Bombus subgenus Psithyrus are closely related to, and resemble, their hosts in looks and size. This common pattern gave rise to the ecological principle "Emery's rule". Others parasitize bees in different families, like Townsendiella, a nomadine apid, two species of which are cleptoparasites of the dasypodaid genus Hesperapis,[78] while the other species in the same genus attacks halictid bees.[79]
Nocturnal bees
Four bee families (Andrenidae, Colletidae, Halictidae, and Apidae) contain some species that are crepuscular. Most are tropical or subtropical, but some live in arid regions at higher latitudes. These bees have greatly enlarged ocelli, which are extremely sensitive to light and dark, though incapable of forming images. Some have refracting superposition compound eyes: these combine the output of many elements of their compound eyes to provide enough light for each retinal photoreceptor. Their ability to fly by night enables them to avoid many predators, and to exploit flowers that produce nectar only or also at night.[80]
Predators, parasites and pathogens

Vertebrate predators of bees include bee-eaters, shrikes and flycatchers, which make short sallies to catch insects in flight.[81] Swifts and swallows[81] fly almost continually, catching insects as they go. The honey buzzard attacks bees' nests and eats the larvae.[82] The greater honeyguide interacts with humans by guiding them to the nests of wild bees. The humans break open the nests and take the honey and the bird feeds on the larvae and the wax.[83] Among mammals, predators such as the badger dig up bumblebee nests and eat both the larvae and any stored food.[84]

Specialist ambush predators of visitors to flowers include crab spiders, which wait on flowering plants for pollinating insects; predatory bugs, and praying mantises,[81] some of which (the flower mantises of the tropics) wait motionless, aggressive mimics camouflaged as flowers.[85] Beewolves are large wasps that habitually attack bees;[81] the ethologist Niko Tinbergen estimated that a single colony of the beewolf Philanthus triangulum might kill several thousand honeybees in a day: all the prey he observed were honeybees.[86] Other predatory insects that sometimes catch bees include robber flies and dragonflies.[81] Honey bees are affected by parasites including acarine and Varroa mites.[87] However, some bees are believed to have a mutualistic relationship with mites.[21]
Relationship with humans
In mythology and folklore

Homer's Hymn to Hermes describes three bee-maidens with the power of divination and thus speaking truth, and identifies the food of the gods as honey. Sources associated the bee maidens with Apollo and, until the 1980s, scholars followed Gottfried Hermann (1806) in incorrectly identifying the bee-maidens with the Thriae.[88] Honey, according to a Greek myth, was discovered by a nymph called Melissa ("Bee"); and honey was offered to the Greek gods from Mycenean times. Bees were also associated with the Delphic oracle and the prophetess was sometimes called a bee.[89]
The image of a community of honey bees has been used from ancient to modern times, in Aristotle and Plato; in Virgil and Seneca; in Erasmus and Shakespeare; Tolstoy, and by political and social theorists such as Bernard Mandeville and Karl Marx as a model for human society.[90] In English folklore, bees would be told of important events in the household, in a custom known as "Telling the bees".[91]
In art and literature

Some of the oldest examples of bees in art are rock paintings in Spain which have been dated to 15,000 BC.[92]
W. B. Yeats's poem The Lake Isle of Innisfree (1888) contains the couplet "Nine bean rows will I have there, a hive for the honey bee, / And live alone in the bee loud glade." At the time he was living in Bedford Park in the West of London.[93] Beatrix Potter's illustrated book The Tale of Mrs Tittlemouse (1910) features Babbity Bumble and her brood (pictured). Kit Williams' treasure hunt book The Bee on the Comb (1984) uses bees and beekeeping as part of its story and puzzle. Sue Monk Kidd's The Secret Life of Bees (2004), and the 2009 film starring Dakota Fanning, tells the story of a girl who escapes her abusive home and finds her way to live with a family of beekeepers, the Boatwrights.
The humorous 2007 animated film Bee Movie used Jerry Seinfeld's first script and was his first work for children; he starred as a bee named Barry B. Benson, alongside Renée Zellweger. Critics found its premise awkward and its delivery tame.[94] Dave Goulson's A Sting in the Tale (2014) describes his efforts to save bumblebees in Britain, as well as much about their biology. The playwright Laline Paull's fantasy The Bees (2015) tells the tale of a hive bee named Flora 717 from hatching onwards.[95]
Beekeeping

Humans have kept honey bee colonies, commonly in hives, for millennia. Beekeepers collect honey, beeswax, propolis, pollen, and royal jelly from hives; bees are also kept to pollinate crops and to produce bees for sale to other beekeepers.
Depictions of humans collecting honey from wild bees date to 15,000 years ago; efforts to domesticate them are shown in Egyptian art around 4,500 years ago.[96] Simple hives and smoke were used;[97][98] jars of honey were found in the tombs of pharaohs such as Tutankhamun. From the 18th century, European understanding of the colonies and biology of bees allowed the construction of the moveable comb hive so that honey could be harvested without destroying the colony.[99][100] Among Classical Era authors, beekeeping with the use of smoke is described in Aristotle's History of Animals Book 9.[101] The account mentions that bees die after stinging; that workers remove corpses from the hive, and guard it; castes including workers and non-working drones, but "kings" rather than queens; predators including toads and bee-eaters; and the waggle dance, with the "irresistible suggestion" of άpοσειονται ("aroseiontai", it waggles) and παρακολουθούσιν ("parakolouthousin", they watch).[102][lower-alpha 2]
Beekeeping is described in detail by Virgil in his Georgics; it is also mentioned in his Aeneid, and in Pliny's Natural History.[102]
As commercial pollinators


Bees play an important role in pollinating flowering plants, and are the major type of pollinator in many ecosystems that contain flowering plants. It is estimated that one third of the human food supply depends on pollination by insects, birds and bats, most of which is accomplished by bees, whether wild or domesticated.[103][104] Over the last half century, there has been a general decline in the species richness of wild bees and other pollinators, probably attributable to stress from increased parasites and disease, the use of pesticides, and a general decrease in the number of wild flowers. Climate change probably exacerbates the problem.[105]
Contract pollination has overtaken the role of honey production for beekeepers in many countries. After the introduction of Varroa mites, feral honey bees declined dramatically in the US, though their numbers have since recovered.[106][107] The number of colonies kept by beekeepers declined slightly, through urbanization, systematic pesticide use, tracheal and Varroa mites, and the closure of beekeeping businesses. In 2006 and 2007 the rate of attrition increased, and was described as colony collapse disorder.[108] In 2010 invertebrate iridescent virus and the fungus Nosema ceranae were shown to be in every killed colony, and deadly in combination.[109][110][111][112] Winter losses increased to about 1/3.[113][114] Varroa mites were thought to be responsible for about half the losses.[115]
Apart from colony collapse disorder, losses outside the US have been attributed to causes including pesticide seed dressings, using neonicotinoids such as Clothianidin, Imidacloprid and Thiamethoxam.[116][117] From 2013 the European Union restricted some pesticides to stop bee populations from declining further.[118] In 2014 the Intergovernmental Panel on Climate Change report warned that bees faced increased risk of extinction because of global warming.[119] In 2018 the European Union decided to ban field use of all three major neonicotinoids; they remain permitted in veterinary, greenhouse, and vehicle transport usage.[120]
Farmers have focused on alternative solutions to mitigate these problems. By raising native plants, they provide food for native bee pollinators like Lasioglossum vierecki[121] and L. leucozonium,[122] leading to less reliance on honey bee populations.
As food producers
Honey is a natural product produced by bees and stored for their own use, but its sweetness has always appealed to humans. Before domestication of bees was even attempted, humans were raiding their nests for their honey. Smoke was often used to subdue the bees and such activities are depicted in rock paintings in Spain dated to 15,000 BC.[92]
Honey bees are used commercially to produce honey.[123] They also produce some substances used as dietary supplements with possible health benefits, pollen,[124] propolis,[125] and royal jelly,[126] though all of these can also cause allergic reactions.
As food

Bees are considered edible insects. People in some countries eat insects, including the larvae and pupae of bees, mostly stingless species. They also gather larvae, pupae and surrounding cells, known as bee brood, for consumption.[127] In the Indonesian dish botok tawon from Central and East Java, bee larvae are eaten as a companion to rice, after being mixed with shredded coconut, wrapped in banana leaves, and steamed.[128][129]

Bee brood (pupae and larvae) although low in calcium, has been found to be high in protein and carbohydrate, and a useful source of phosphorus, magnesium, potassium, and trace minerals iron, zinc, copper, and selenium. In addition, while bee brood was high in fat, it contained no fat soluble vitamins (such as A, D, and E) but it was a good source of most of the water-soluble B-vitamins including choline as well as vitamin C. The fat was composed mostly of saturated and monounsaturated fatty acids with 2.0% being polyunsaturated fatty acids.[130][131]
As alternative medicine
Apitherapy is a branch of alternative medicine that uses honey bee products, including raw honey, royal jelly, pollen, propolis, beeswax and apitoxin (Bee venom).[132] The claim that apitherapy treats cancer, which some proponents of apitherapy make, remains unsupported by evidence-based medicine.[133][134]
Stings
The painful stings of bees are mostly associated with the poison gland and the Dufour's gland which are abdominal exocrine glands containing various chemicals. In Lasioglossum leucozonium, the Dufour's Gland mostly contains octadecanolide as well as some eicosanolide. There is also evidence of n-triscosane, n-heptacosane,[135] and 22-docosanolide.[136] However, the secretions of these glands could also be used for nest construction.[135]
See also
Notes
- Triassic nests in a petrified forest in Arizona, implying that bees evolved much earlier,[7] are now thought to be beetle borings.[8]
- In D'Arcy Thompson's translation: "At early dawn they make no noise, until some one particular bee makes a buzzing noise two or three times and thereby awakes the rest; hereupon they all fly in a body to work. By and by they return and at first are noisy; ... until at last some one bee flies round about, making a buzzing noise, and apparently calling on the others to go to sleep".[101]
References
- Danforth, B. N.; Sipes, S.; Fang, J.; Brady, S. G. (October 2006). "The history of early bee diversification based on five genes plus morphology". PNAS. 103 (41): 15118–15123. Bibcode:2006PNAS..10315118D. doi:10.1073/pnas.0604033103. PMC 1586180. PMID 17015826.
- Michener, Charles D. (2000). The Bees of the World. Johns Hopkins University Press. pp. 19–25. ISBN 0-8018--6133-0.
- "Widespread losses of pollinating insects revealed across Britain". The Guardian. 26 March 2019.
- Cardinal, Sophie; Danforth, Bryan N. (2011). "The Antiquity and Evolutionary History of Social Behavior in Bees". PLOS One. 6 (6): e21086. Bibcode:2011PLoSO...621086C. doi:10.1371/journal.pone.0021086. PMC 3113908. PMID 21695157.
- Poinar, G. O.; Danforth, B. N. (2006). "A fossil bee from Early Cretaceous Burmese amber" (PDF). Science. 314 (5799): 614. doi:10.1126/science.1134103. PMID 17068254. S2CID 28047407. Archived from the original (PDF) on 4 December 2012.
- Engel, Michael S. (2001). "Monophyly and Extensive Extinction of Advanced Eusocial Bees: Insights from an Unexpected Eocene Diversity". PNAS. National Academy of Sciences. 98 (4): 1661–1664. Bibcode:2001PNAS...98.1661E. doi:10.1073/pnas.041600198. JSTOR 3054932. PMC 29313. PMID 11172007.
- Buchmann, Stephen L.; Nabhan, Gary Paul (2012). The Forgotten Pollinators. Island Press. pp. 41–42. ISBN 978-1-59726-908-7. Archived from the original on 27 May 2016.
- Lucas, Spencer G.; Minter, Nicholas J.; Hunt, Adrian P. (February 2010). "Re-evaluation of alleged bees' nests from the Upper Triassic of Arizona". Palaeogeography, Palaeoclimatology, Palaeoecology. 286 (3–4): 194–201. Bibcode:2010PPP...286..194L. doi:10.1016/j.palaeo.2010.01.010.
- Danforth, Bryan; Cardinal, Sophie; Praz, Christophe; Almeida, Eduardo; Michez, Denis (28 August 2012). "The Impact of Molecular Data on Our Understanding of Bee Phylogeny and Evolution". Annual Review of Entomology. 58: 57–78. doi:10.1146/annurev-ento-120811-153633. PMID 22934982. S2CID 28274420.
- Almeida, Eduardo A. B.; Pie, Marcio R.; Brady, Sean G.; Danforth, Bryan N. (2012). "Biogeography and diversification of colletid bees (Hymenoptera: Colletidae): emerging patterns from the southern end of the world" (PDF). Journal of Biogeography. 39 (3): 526–544. doi:10.1111/j.1365-2699.2011.02624.x. Archived (PDF) from the original on 21 September 2013.
- Michez, Denis; Nel, Andre; Menier, Jean-Jacques; Rasmont, Pierre (2007). "The oldest fossil of a melittid bee (Hymenoptera: Apiformes) from the early Eocene of Oise (France)" (PDF). Zoological Journal of the Linnean Society. 150 (4): 701–709. doi:10.1111/j.1096-3642.2007.00307.x. Archived (PDF) from the original on 23 September 2015.
- Sarzetti, Laura C.; Lanandeira, Conrad C.; Genise, Jorge F. (2008). "A Leafcutter Bee Trace Fossil from the Middle Eocene of Patagonia, Argentina, and a Review of Megachilid (Hymenoptera) Ichnology". Palaeontology. 51 (4): 933–994. doi:10.1111/j.1475-4983.2008.00787.x. Archived (PDF) from the original on 24 September 2015.
- Dewulf, Alexandre; De Meulemeester, Thibaut; Dehon, Manuel; Engel, Michael S.; Michez, Denis (2014). "A new interpretation of the bee fossil Melitta willardi Cockerell (Hymenoptera, Melittidae) based on geometric morphometrics of the wing". ZooKeys (389): 35–48. doi:10.3897/zookeys.389.7076. PMC 3974431. PMID 24715773.
- Engel, M.S.; Archibald, S.B. (2003). "An Early Eocene bee (Hymenoptera: Halictidae) from Quilchena, British Columbia" (PDF). The Canadian Entomologist. 135 (1): 63–69. doi:10.4039/n02-030. hdl:1808/16473.
- Engel, M.S. (1995). "Neocorynura electra, a New Fossil Bee Species from Dominican Amber (Hymenoptera:Halictidae)". Journal of the New York Entomological Society. 103 (3): 317–323. JSTOR 25010174.
- Engel, M.S. (2000). "Classification of the bee tribe Augochlorini (Hymenoptera, Halictidae)" (PDF). Bulletin of the American Museum of Natural History. 250: 1. doi:10.1206/0003-0090(2000)250<0001:COTBTA>2.0.CO;2. hdl:2246/1598. Archived (PDF) from the original on 10 January 2011.
- Houston, T.F. (1987). "Fossil brood cells of stenotritid bees (Hymenoptera: Apoidea) from the Pleistocene of South Australia". Transactions of the Royal Society of South Australia. 1111–2: 93–97. Archived from the original on 1 July 2015.
- Armbruster, W. Scott (2012). "3". In Patiny, Sébastien (ed.). Evolution of Plant-Pollinator Relationships. Cambridge University Press. pp. 45–67.
- Michener, Charles Duncan (1974). The Social Behavior of the Bees: A Comparative Study. Harvard University Press. pp. 22–78. ISBN 978-0-674-81175-1.
- Biani, Natalia B.; Mueller, Ulrich G.; Wcislo, William T. (June 2009). "Cleaner Mites: Sanitary Mutualism in the Miniature Ecosystem of Neotropical Bee Nests" (PDF). The American Naturalist. 173 (6): 841–847. doi:10.1086/598497. hdl:2152/31261. PMID 19371167.
- Klimov, Pavel B.; OConnor, Barry M.; Knowles, L. Lacey (June 2007). "Museum Specimens And Phylogenies Elucidate Ecology's Role in Coevolutionary Associations Between Mites And Their Bee Hosts" (PDF). Evolution. 61 (6): 1368–1379. doi:10.1111/j.1558-5646.2007.00119.x. hdl:2027.42/74970. PMID 17542846. S2CID 32318137.
- Debevec, Andrew H.; Cardinal, Sophie; Danforth, Bryan N. (2012). "Identifying the sister group to the bees: a molecular phylogeny of Aculeata with an emphasis on the superfamily Apoidea" (PDF). Zoologica Scripta. 41 (5): 527–535. doi:10.1111/j.1463-6409.2012.00549.x. Archived (PDF) from the original on 23 September 2015.
- Hedtke, Shannon M.; Patiny, Sébastien; Danforth, Bryan M. (2013). "The bee tree of life: a supermatrix approach to apoid phylogeny and biogeography". BMC Evolutionary Biology. 13 (138): 138. doi:10.1186/1471-2148-13-138. PMC 3706286. PMID 23822725.
- Grimaldi, David; Engel, Michael S. (2005). Evolution of the Insects. Cambridge University Press. p. 454. ISBN 978-0-521-82149-0. Archived from the original on 28 March 2018.
- "Anatomy of the Honey Bee". Extension. 19 June 2014. Archived from the original on 1 July 2015. Retrieved 30 June 2015.
- Messer, A. C. (1984). "Chalicodoma pluto: The World's Largest Bee Rediscovered Living Communally in Termite Nests (Hymenoptera: Megachilidae)". Journal of the Kansas Entomological Society. 57 (1): 165–168. JSTOR 25084498.
- Sakagami, Shôichi F.; Zucchi, Ronaldo (1974). "Oviposition Behavior of Two Dwarf Stingless Bees, Hypotrigona (Leurotrigona) muelleri and H. (Trigonisca) duckei, with Notes on the Temporal Articulation of Oviposition Process in Stingless Bees" (PDF). Journal of the Faculty of Science Hokkaido University Series Vi. Zoology. 19 (2): 361–421. Archived (PDF) from the original on 4 March 2016.
- Hughes, W. O. H.; Oldroyd, B. P.; Beekman, M.; Ratnieks, F. L. W. (2008). "Ancestral Monogamy Shows Kin Selection is Key to the Evolution of Eusociality". Science. 320 (5880): 1213–1216. Bibcode:2008Sci...320.1213H. doi:10.1126/science.1156108. PMID 18511689. S2CID 20388889.
- Hamilton, W. D. (20 March 1964). "The Genetical Evolution of Social Behaviour II". Journal of Theoretical Biology. 7 (1): 17–52. doi:10.1016/0022-5193(64)90039-6. PMID 5875340.
- Hughes, William O. H.; Oldroyd, Benjamin P.; Beekman, Madeleine; Ratnieks, Francis L. W. (May 2008). "Ancestral Monogamy Shows Kin Selection Is Key to the Evolution of Eusociality". Science. American Association for the Advancement of Science. 320 (5880): 1213–1216. Bibcode:2008Sci...320.1213H. doi:10.1126/science.1156108. PMID 18511689. S2CID 20388889.
- Gullan, P. J.; Cranston, P. S. (2014). The Insects: An Outline of Entomology (5th ed.). Wiley Blackwell. pp. 328, 348–350. ISBN 978-1-118-84615-5.
- Nowak, Martin; Tarnita, Corina; Wilson, E.O. (2010). "The evolution of eusociality". Nature. 466 (7310): 1057–1062. Bibcode:2010Natur.466.1057N. doi:10.1038/nature09205. PMC 3279739. PMID 20740005.
- Brady, Seán G.; Sipes, Sedonia; Pearson, Adam; Danforth, Bryan N. (2006). "Recent and simultaneous origins of eusociality in halictid bees". Proceedings of the Royal Society of London B: Biological Sciences. 273 (1594): 1643–1649. doi:10.1098/rspb.2006.3496. ISSN 0962-8452. PMC 1634925. PMID 16769636.
- Wilson, Edward O (1971). The Insect Societies. Cambridge, Mass: Belknap Press of Harvard University Press.
- Sanford, Malcolm T. (2006). "The Africanized Honey Bee in the Americas: A Biological Revolution with Human Cultural Implications". Apis Enterprises. Archived from the original on 29 March 2015. Retrieved 29 March 2015.
- Roubik, D. W. (2006). "Stingless bee nesting biology" (PDF). Apidologie. 37 (2): 124–143. doi:10.1051/apido:2006026.
- "Bumblebee nests". Bumblebee Conservation Trust. Archived from the original on 22 September 2017. Retrieved 26 June 2015.
- "Bumblebee Specialist Group: 2011 Update" (PDF). IUCN. Archived (PDF) from the original on 3 December 2012. Retrieved 7 October 2012.
- Brooks, R. W.; Roubik, D. W. (1983). "A Halictine bee with distinct castes: Halictus hesperus (Hymenoptera: Halictidae) and its bionomics in Central Panama". Sociobiology. 7: 263–282.
- Eickwort, G. C.; Eickwort, J. M.; Gordon, J.; Eickwort, M. A.; Wcislo, W. T. (1996). "Solitary behavior in a high-altitude population of the social sweat bee Halictus rubicundus (Hymenoptera: Halictidae)". Behavioral Ecology and Sociobiology. 38 (4): 227–233. doi:10.1007/s002650050236. S2CID 12868253.
- Yanega, D. (1993). "Environmental effects on male production and social structure in Halictus rubicundus (Hymenoptera: Halictidae)". Insectes Sociaux. 40: 169–180. doi:10.1007/bf01240705. S2CID 44934383.
- Michener, Charles Duncan (1974). The Social Behavior of the Bees: A Comparative Study. Harvard University Press. p. 308. ISBN 978-0-674-81175-1. Archived from the original on 24 December 2016.
- Parker, Frank D.; Torchio, Philip F. (1 October 1980). "Management of Wild Bees". Beesource Beekeeping Community. Archived from the original on 26 June 2015. Retrieved 26 June 2015.
- "Solitary Bees (Hymenoptera)". Royal Entomological Society. Archived from the original on 2 July 2017. Retrieved 12 October 2015.
- "Other bees". Bumblebee Conservation Trust. Archived from the original on 5 September 2015. Retrieved 12 October 2015.
- Westreich, Lila. "Spring signals female bees to lay the next generation of pollinators". The Conversation. Retrieved 8 October 2020.
- Eickwort, George C. (1975). "Gregarious Nesting of the Mason Bee Hoplitis anthocopoides and the Evolution of Parasitism and Sociality Among Megachilid Bees". Evolution. 29 (1): 142–150. doi:10.2307/2407147. JSTOR 2407147. PMID 28563288.
- Alcock, John (1 May 1999). "The Nesting Behavior of Dawson's Burrowing Bee, Amegilla dawsoni (Hymenoptera: Anthophorini), and the Production of Offspring of Different Sizes". Journal of Insect Behavior. 12 (3): 363–384. doi:10.1023/A:1020843606530. ISSN 0892-7553. S2CID 24832889.
- Roubik, David W. (1992). Ecology and Natural History of Tropical Bees. Cambridge University Press. p. 15. ISBN 978-0-521-42909-2. Archived from the original on 17 June 2016.
- "The bumblebee lifecycle". Bumblebee Conservation Trust. Archived from the original on 29 June 2015. Retrieved 1 July 2015.
- "Learning About Honey Bees". The South Carolina Mid-State Beekeepers Association. Archived from the original on 1 July 2015. Retrieved 1 July 2015.
- "Solitary Bees". National Bee Unit. Archived from the original on 1 July 2015. Retrieved 1 July 2015.
- Shuckard, William Edward (1866). British bees: an introduction to the study of the natural history and economy of the bees indigenous to the British Isles. L. Reeve & Co. pp. 18–23.
- Ingram, Jay (2001) The Barmaid's Brain, Aurum Press, pp. 91–92, ISBN 0716741202.
- Adams, Cecil (4 May 1990). "Is it aerodynamically impossible for bumblebees to fly?". The Straight Dope. Archived from the original on 3 March 2009. Retrieved 7 March 2009.
- "Life, animal and plant news, articles and features". New Scientist. 9 March 2016. Archived from the original on 7 October 2008. Retrieved 16 March 2016.
- "Images of flight". New Scientist. Archived from the original on 23 March 2016. Retrieved 16 March 2016.
- "Deciphering the Mystery of Bee Flight". California Institute of Technology. 29 November 2005. Archived from the original on 17 September 2016. Retrieved 8 September 2016. Re: work of Dr. Michael H. Dickinson.
- Altshuler, Douglas L.; Dickson, William B.; Vance, Jason T.; Roberts, Stephen P.; Dickinson, Michael H. (2005). "Short-amplitude high-frequency wing strokes determine the aerodynamics of honeybee flight". Proceedings of the National Academy of Sciences. 102 (50): 18213–18218. Bibcode:2005PNAS..10218213A. doi:10.1073/pnas.0506590102. PMC 1312389. PMID 16330767.
- von Frisch, Karl (1953). The Dancing Bees. Harcourt, Brace & World. pp. 93–96.
- Menzel, Randolf; Greggers, Uwe; Smith, Alan; Berger, Sandra; Brandt, Robert; Brunke, Sascha; Bundrock, Gesine; Hülse, Sandra; Plümpe, Tobias; Schaupp, Schaupp; Schüttler, Elke; Stach, Silke; Stindt, Jan; Stollhoff, Nicola; Watzl, Sebastian (2005). "Honey bees Navigate According to a Map-Like Spatial Memory". PNAS. 102 (8): 3040–3045. Bibcode:2005PNAS..102.3040M. doi:10.1073/pnas.0408550102. PMC 549458. PMID 15710880.
- "How honey bee gut bacteria help to digest their pollen-rich diet". ScienceDaily. Retrieved 2 January 2020.
- "Bee gut microbes have a division of labor when it comes to metabolizing complex polysaccharides". phys.org. Retrieved 2 January 2020.
- Zheng, Hao; Perreau, Julie; Powell, J. Elijah; Han, Benfeng; Zhang, Zijing; Kwong, Waldan K.; Tringe, Susannah G.; Moran, Nancy A. (December 2019). "Division of labor in honey bee gut microbiota for plant polysaccharide digestion". Proceedings of the National Academy of Sciences. 116 (51): 25909–25916. doi:10.1073/pnas.1916224116. ISSN 0027-8424. PMC 6926048. PMID 31776248.
- Mateus, Sidnei; Noll, Fernando B. (February 2004). "Predatory behavior in a necrophagous bee Trigona hypogea (Hymenoptera; Apidae, Meliponini)". Naturwissenschaften. 91 (2): 94–96. Bibcode:2004NW.....91...94M. doi:10.1007/s00114-003-0497-1. ISSN 1432-1904. PMID 14991148. S2CID 26518321.
- Waser, Nickolas M. (2006). Plant-Pollinator Interactions: From Specialization to Generalization. University of Chicago Press. pp. 110–. ISBN 978-0-226-87400-5. Archived from the original on 28 March 2018.
- Dafni, Amots; Hesse, Michael; Pacini, Ettore (2012). Pollen and Pollination. Springer Science & Business Media. p. 80. ISBN 978-3-7091-6306-1. Archived from the original on 28 March 2018.
- Suttona, Gregory P.; Clarkea, Dominic; Morleya, Erica L.; Robert, Daniel (2016). "Mechanosensory hairs in bumblebees (Bombus terrestris) detect weak electric fields". PNAS. 113 (26): 7261–7265. doi:10.1073/pnas.1601624113. PMC 4932954. PMID 27247399.
- Muth, Felicity; Francis, Jacob S.; Leonard, Anne S. (2016). "Bees use the taste of pollen to determine which flowers to visit". Biology Letters. 12 (7): 20160356. doi:10.1098/rsbl.2016.0356. PMC 4971173. PMID 27405383.
- Hurd, P.D. Jr.; Linsley, E.G. (1975). "The principal Larrea bees of the southwestern United States". Smithsonian Contributions to Zoology. 193 (193): 1–74. doi:10.5479/si.00810282.193.
- Thorp, Robbin W.; Horning, Donald S.; Dunning, Lorry L. (1983). Bumble Bees and Cuckoo Bumble Bees of California (Hymenoptera, Apidae). University of California Press. p. 9. ISBN 978-0-520-09645-5. Archived from the original on 5 January 2017.
Of the forms of mimicry, two relate to Bombini. Batesian mimicry .. is exemplified by members of several families of flies: Syrphidae, Asilidae, Tabanidae, Oestridae, and Bombyliidae (Gabritschevsky, 1926).
- Cott, Hugh (1940). Adaptive Coloration in Animals. Oxford University Press. pp. 196, 403 and passim.
- "Bee Orchids and Insect Mimicry". Natural History Museum. Archived from the original on 8 July 2015. Retrieved 1 July 2015.
- "Obligate Brood Parasitism". Aculeata Research Group. Archived from the original on 7 July 2015. Retrieved 30 June 2015.
- "Brood Parasitism". Amateur Entomologists' Society. Archived from the original on 2 July 2015. Retrieved 30 June 2015.
- Gjershaug, Jan Ove (5 June 2009). "The social parasite bumblebee Bombus hyperboreus Schönherr, 1809 usurp nest of Bombus balteatus Dahlbom, 1832 (Hymenoptera, Apidae) in Norway" (PDF). Norwegian Journal of Entomology 56(1): 28–31. Retrieved 26 September 2015.
- Gullan, P. J.; Cranston, P. S. (2014). The Insects: An Outline of Entomology (5th ed.). Wiley Blackwell. p. 347. ISBN 978-1-118-84615-5.
- Rozen, Jerome George; McGinley, Ronald J. (1991). "Biology and Larvae of the Cleptoparasitic Bee Townsendiella pulchra and Nesting Biology of its Host Hesperapis larreae (Hymenoptera, Apoidea)". American Museum Novitates. 3005. hdl:2246/5032.
- Moure, Jesus S.; Hurd, Paul David (1987). An Annotated Catalog of the Halictid Bees of the Western Hemisphere (Hymenoptera, Halictidae). Smithsonian Institution Press. pp. 28–29.
- Warrant, Eric J. (June 2008). "Seeing in the dark: vision and visual behaviour in nocturnal bees and wasps". Journal of Experimental Biology. 211 (11): 1737–1746. doi:10.1242/jeb.015396. PMID 18490389.
- Chittka, Lars; Thomson, James D. (28 May 2001). Cognitive Ecology of Pollination: Animal Behaviour and Floral Evolution. Cambridge University Press. pp. 215–216. ISBN 978-1-139-43004-3. Archived from the original on 24 December 2016.
- "Hornet attacks kill dozens in China". The Guardian. 26 September 2013. Archived from the original on 6 September 2015. Retrieved 18 June 2015.
- Friedmann, Herbert (1955). "The Honey-Guides". Bulletin of the United States National Museum (208): 1–292. doi:10.5479/si.03629236.208.1. hdl:10088/10101.
- "What predators do bumblebees have?". Bumblebee Conservation Trust. Archived from the original on 29 June 2015. Retrieved 29 June 2015.
- Choi, Charles Q. (30 November 2013). "Found! First Known Predator To Lure Prey By Mimicking Flowers". LiveScience. Archived from the original on 30 June 2015. Retrieved 2 July 2015.
the color of the orchid mantis was indistinguishable from 13 species of wild flowers in the areas the predator lived. ... The orchid mantis is unique in that the mantis itself is the attractive stimulus.
- Tinbergen, Niko (1958). Curious Naturalists. Methuen. p. 21.
- "Honey Bee Disorders: Honey Bee Parasites". University of Georgia. Archived from the original on 1 July 2015. Retrieved 29 June 2015.
- Susan Scheinberg, "The Bee Maidens of the Homeric Hymn to Hermes", in Albert Heinrichs, ed., Harvard Studies in Classical Philology (Cambridge MA: Harvard University Press, 1980), 11. ISBN 0674379306; and many others since questioning Gottfried Hermann's 1806 equation of the Thriae with bee-maidens. Heinrich Gottfried, Homeri nomine dignissimum/Homeric Hymns (Leipzig: 1806), 346 and cxiii. Many took Hermann's unfounded identification at face-value, repeating it ad nauseam, e.g. Hilda M. Ransome, The Sacred Bee in Ancient Times and Folklore (NY: Courier, 1937; reprinted as recently as NY: Dover, 2012), 97. ISBN 0486122980
- Scheinberg, Susan (1979). "The Bee Maidens of the Homeric Hymn to Hermes". Harvard Studies in Classical Philology. 83: 1–28. doi:10.2307/311093. JSTOR 311093.
- Wilson, Bee (2004). The Hive: the Story of the Honeybee. London: John Murray. ISBN 0-7195-6598-7.
- Steve Roud (6 April 2006). The Penguin Guide to the Superstitions of Britain and Ireland. Penguin Books. p. 128. ISBN 978-0-14-194162-2. Archived from the original on 28 November 2016.
- Encyclopaedia of the History of Science, Technology, and Medicine in Non-Western Cultures. Springer Science & Business Media. 12 March 2008. p. 1074. ISBN 978-1-4020-4559-2. Archived from the original on 4 July 2014.
- Deering, Chris. "Yeats in Bedford Park". ChiswickW4.com. Archived from the original on 30 June 2015. Retrieved 28 June 2015.
- "Bee Movie". Rotten Tomatoes. Archived from the original on 23 June 2015. Retrieved 30 June 2015.
- Jones, Gwyneth (21 May 2014). "The Bees by Laline Paull review – a fantasy with a sting in its tail". The Guardian. Archived from the original on 1 July 2015. Retrieved 28 June 2015.
- "Ancient Egypt: Bee-keeping". Reshafim.org.il. 6 April 2003. Archived from the original on 9 March 2016. Retrieved 16 March 2016.
- "Beekeeping in Ancient Egypt". Bee Lore. 23 February 2008. Archived from the original on 22 March 2016. Retrieved 16 March 2016.
- Bodenheimer, F. S. (1960). Animal and Man in Bible Lands. Brill Archive. p. 79.
- Thomas Wildman, A Treatise on the Management of Bees (London, 1768, 2nd edn 1770).
- Harissis, H. V.; Mavrofridis, G. (2012). "A 17th Century Testimony on the Use of Ceramic Top-bar Hives". Bee World. 89 (3): 56–57. doi:10.1080/0005772x.2012.11417481. S2CID 85120138. Archived from the original on 19 October 2015.
- Aristotle; Thompson, D'Arcy (trans.) (1910). The Works of Aristotle. Clarendon Press. pp. Book 9, Section 40.
- Whitfield, B. G. (October 1956). "Cambridge University Press and The Classical Association are collaborating with JSTOR to digitize, preserve and extend access to Greece & Rome. Virgil and the Bees: A Study in Ancient Apicultural Lor". Greece and Rome. 3 (2): 99–117. doi:10.1017/S0017383500015126. JSTOR 641360.
- Yang, Sarah (25 October 2006). "Pollinators help one-third of world's crop production, says new study". UC Berkeley. Archived from the original on 9 July 2015. Retrieved 29 June 2015.
- Connor, Steve (16 June 2015). "Wild bees just as important as domesticated bees for pollinating food crops". The Independent. Archived from the original on 6 September 2017.
Wild bees have become as important as domesticated honeybees in pollinating food crops around the world due to the dramatic decline in number of healthy honeybee colonies over the past half century, a study has found.
- Goulson, Dave; Nicholls, Elizabeth; Botías, Cristina; Rotheray, Ellen L. (2015). "Bee declines driven by combined stress from parasites, pesticides, and lack of flowers". Science. 347 (6229): 1255957. doi:10.1126/science.1255957. PMID 25721506. S2CID 206558985.CS1 maint: multiple names: authors list (link)
- Loper, Gerald M.; Sammataro, Diana; Finley, Jennifer; Cole, Jerry (2006). "Feral honey bees in southern Arizona, 10 years after varroa infestation". American Bee Journal. 146: 521–524.
- Rangel, Juliana; Giresi, Melissa; Pinto, Maria Alice; Baum, Kristen A.; Rubink, William L.; Coulson, Robert N.; Johnston, John Spencer (2016). "Africanization of a feral honey bee (Apis mellifera) population in South Texas: does a decade make a difference?". Ecology and Evolution. 6 (7): 2158–2169. doi:10.1002/ece3.1974. PMC 4782243. PMID 27069571.
- "Honey Bee Die-Off Alarms Beekeepers, Crop Growers and Researchers". Pennsylvania State University College of Agricultural Sciences. 29 January 2007. Archived from the original on 17 May 2008.
- Johnson, Kirk (6 October 2010) Scientists and Soldiers Solve a Bee Mystery Archived 7 October 2010 at the Wayback Machine. The New York Times.
- Eban, Katherine (8 October 2010). "What a scientist didn't tell the New York Times about his study on bee deaths". CNN. Archived from the original on 19 October 2012. Retrieved 20 August 2012.
- Jerry J. Bromenshenk; Colin B. Henderson; Charles H. Wick; Michael F. Stanford; Alan W. Zulich; Rabih E. Jabbour; Samir V. Deshpande; Patrick E. McCubbin; Robert A. Seccomb; Phillip M. Welch; Trevor Williams; David R. Firth; Evan Skowronski; Margaret M. Lehmann; Shan L. Bilimoria; Joanna Gress; Kevin W. Wanner; Robert A. Cramer Jr (6 October 2010). "Iridovirus and Microsporidian Linked to Honey Bee Colony Decline". PLOS One. 5 (10): e13181. Bibcode:2010PLoSO...513181B. doi:10.1371/journal.pone.0013181. PMC 2950847. PMID 20949138.
- "Honey bees in US facing extinction" Archived 6 September 2008 at the Wayback Machine, The Daily Telegraph (London), 14 March 2007.
- Benjamin, Alison (2 May 2010) Fears for crops as shock figures from America show scale of bee catastrophe Archived 4 December 2013 at the Wayback Machine. The Observer (London).
- "Beekeepers Report Continued Heavy Losses From Colony Collapse Disorder". Sciencedaily.com. 12 May 2008. Archived from the original on 31 July 2010. Retrieved 22 June 2010.
- "Hiver fatal pour la moitié des colonies d'abeilles en Suisse". Radio Télévision Suisse. 22 May 2012. Archived from the original on 12 November 2012. Retrieved 22 May 2012.
- Storkstad, Erik (30 March 2012). "Field Research on Bees Raises Concern About Low-Dose Pesticides". Science. 335 (6076): 1555. Bibcode:2012Sci...335.1555S. doi:10.1126/science.335.6076.1555. PMID 22461580. S2CID 206597443.
- "EFSA identifies risks to bees from neonicotinoids | European Food Safety Authority". Efsa.europa.eu. 20 September 2012. Archived from the original on 28 July 2015. Retrieved 16 March 2016.
- "EU moves to protect bees". 3 News NZ. 30 April 2013. Archived from the original on 29 July 2013.
- Gosden, Emily (29 March 2014) Bees and the crops they pollinate are at risk from climate change, IPCC report to warn Archived 29 August 2014 at the Wayback Machine The Daily Telegraph (London). Retrieved 30 March 2014
- Carrington, Damian (27 April 2018). "EU agrees total ban on bee-harming pesticides". The Guardian.
- Kuehn, F. Coordinator. (2015). Farming for native bees. World Wide Web electronic publication. Retrieved from "Archived copy". Archived from the original on 30 September 2015. Retrieved 4 November 2015.CS1 maint: archived copy as title (link). (Accessed: 22 September 2015).
- Adamson, Nancy Lee. An Assessment of Non-Apis Bees as Fruit and Vegetable Crop Pollinators in Southwest Virginia Archived 20 November 2015 at the Wayback Machine. Diss. 2011. Web. 15 October 2015.
- Hunt, C.L.; Atwater, H.W. (7 April 1915). Honey and Its Uses in the Home. US Department of Agriculture, Farmers' Bulletin, No. 653. Retrieved 14 July 2015.CS1 maint: uses authors parameter (link)
- Sanford, Malcolm T. "Producing Pollen". University of Florida, Institute of Food and Agricultural Sciences. Archived from the original on 13 January 2007. Retrieved 15 July 2015.
- "Propolis:MedlinePlus Supplements". U.S. National Library of Medicine. 19 January 2012. Archived from the original on 30 June 2016.
- European Food Safety Authority (EFSA) Panel on Dietetic Products, Nutrition and Allergies (2011). "Scientific Opinion". EFSA Journal. 9 (4): 2083. doi:10.2903/j.efsa.2011.2083.
- Holland, Jennifer (14 May 2013). "U.N. Urges Eating Insects: 8 Popular Bugs to Try". National Geographic. Archived from the original on 16 July 2015. Retrieved 16 July 2015.
- "Botok Tempe Tahu Teri (Botok Tempe Tofu Anchovy)". Tasty Indonesian Food. Archived from the original on 26 June 2015. Retrieved 22 June 2015. (This particular Botok recipe uses anchovies, not bees)
- Haris, Emmaria (6 December 2013). "Sensasi Rasa Unik Botok Lebah yang Menyengat (Unique taste sensation botok with stinging bees)" (in Indonesian). Sayangi.com. Archived from the original on 22 June 2015. Retrieved 22 June 2015.
- Fink, Mark D. (2007). "Nutrient Composition of Bee Brood and its Potential as Human Food". Ecology of Food and Nutrition. Taylor & Francis, Ecology Food and Nutrition. 44 (4): 257–270. doi:10.1080/03670240500187278. S2CID 84191573.
- Annette Bruun Jensen (2016). "Standard methods for Apis mellifera brood as human food". Journal of Apicultural Research. Taylor & Francis, Journal of Apicultural Research. 58 (2): 1–28. doi:10.1080/00218839.2016.1226606.
- "What is apitherapy?". MedicineWorld.Org. Archived from the original on 18 June 2015. Retrieved 20 January 2018.
- Barry R., Cassileth (2011). "Chapter 36: Apitherapy". The Complete Guide to Complementary Therapies in Cancer Care: Essential Information for Patients, Survivors and Health Professionals. World Scientific. pp. 221–224. ISBN 978-981-4335-66-9. Archived from the original on 7 March 2017.
- Ades, Terri B.; Russel, Jill, eds. (2009). "Chapter 9: Pharmacologic and Biologic Therapies". American Cancer Society Complete Guide to Complementary and Alternative Cancer Therapies (2nd ed.). American Cancer Society. pp. 704–708. ISBN 978-0-944235-71-3.
- Hefetz, Abraham; Blum, Murray; Eickwort, George; Wheeler, James (1978). "Chemistry of the dufour's gland secretion of halictine bees". Comparative Biochemistry and Physiology B. 61 (1): 129–132. doi:10.1016/0305-0491(78)90229-8.
- Johansson, Ingela (1982). "Systematic relationship of halictinae bees based on the pattern of macrocyclic lactones in the Dufour gland secretion". Insect Biochemistry. 12 (2): 161–170. doi:10.1016/0020-1790(82)90004-X.
External links
| Wikiquote has quotations related to: Bees |
| Wikimedia Commons has media related to Anthophila (Bees). |
| Wikispecies has information related to Apoidea. |
| Wikibooks has a book on the topic of: Beekeeping/Solitary_Bee |
- "Bees". Encyclopedia of Life.
- "Apoidea" at All Living Things—images, identification guides, and maps of bees
- Bee Genera of the World
- Anthophila (Apoidea)—Bees—North American species of bees at BugGuide
- Native Bees of North America at BugGuide
- "Bee declines driven by combined stress from parasites, pesticides, and lack of flowers"—Science
| Officials | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Types | |||||||||||||||||
| Charges of heraldic achievem -ents (List) See also: Canting Alliance (Courtesy) Funerary |
| ||||||||||||||||
| Tinctures Rules Tricking Hatching Fimbriation (with black and white rendering) |
| ||||||||||||||||
| External | |||||||||||||||||
| Applications | |||||||||||||||||
| See also | |||||||||||||||||
| |||||||||||||||||
| Authority control |
|---|