Dinocephalosaurus
Dinocephalosaurus (meaning "terrible-headed reptile") is a genus of long necked, aquatic protorosaur that inhabited the Triassic seas of China. The genus contains the type and only known species, D. orientalis, which was named by Li in 2003. Unlike other long-necked protorosaurs (which form a group known as the tanystropheids), Dinocephalosaurus convergently evolved a long neck not through elongation of individual cervical vertebrae, but through the addition of cervical vertebrae that each have a moderate length. Like other tanystropheids, however, Dinocephalosaurus probably used its long neck to hunt for prey, utilizing a combination of suction, created by the expansion of the throat, and the fang-like teeth of the jaws to ensnare prey. It was probably a marine animal by necessity, as suggested by the poorly-ossified and paddle-like limbs which would have prevented it from going ashore.
| Dinocephalosaurus | |
|---|---|
 | |
| Photograph (a) of specimen LPV 30280, which contains an fetus (c) and a perleidid fish (e) | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Family: | †Tanystropheidae |
| Genus: | †Dinocephalosaurus Li, 2003 |
| Species: | †D. orientalis |
| Binomial name | |
| †Dinocephalosaurus orientalis Li, 2003 | |
Specimens belonging to the genus were first discovered in a locality near Xinmin in Guizhou, China in 2002. At the same locality, which dates to 244 million years ago, other marine reptiles such as Mixosaurus, Keichousaurus, and Wumengosaurus have also been found. While the type specimen consisted only of a skull and the very front of the neck, additional specimens soon revealed the complete form of the body. Further discoveries of Dinocephalosaurus specimens were made in Luoping, Yunnan, China, starting in 2008. At this locality, Dinocephalosaurus would have lived alongside Mixosaurus, Dianopachysaurus, and Sinosaurosphargis. One specimen discovered at the Luoping locality preserves a fetus within its abdomen, indicating that Dinocephalosaurus gave birth to live young like many other marine reptiles. Dinocephalosaurus is the only known member of the Archosauromorpha to give live birth, with the possible exception of the metriorhynchids, a group of marine crocodylomorphs.
Description

Dinocephalosaurus was a large member of the Protorosauria, attaining a maximum body length of at least 3.5 m (11 ft),[1] compared to a maximum of 3 metres (9.8 ft) for Tanystropheus.[2][3] The known specimens are probably mature, given that they have fused skull bones and lack the multi-cusped teeth seen in juvenile Tanystropheus.[4] Like Tanystropheus, Dinocephalosaurus has an exceptionally long neck (1.7 m (5 ft 7 in) long) relative to its torso (1 m (3 ft 3 in) excluding the tail).[1]
Skull
The skull of Dinocephalosaurus is low and narrow, with a long premaxilla and maxilla compared to those of Tanystropheus. Both the premaxilla and the maxilla meet at the anteroventral (front bottom) corner and contribute to the border of the nostril, which is located at the anterior (front) end of a long recess that extends along the snout in front of the eye socket (the antorbital depression). The bottom margins of the two snout bones are respectively lined with five and twelve long and nail-like teeth; the third premaxillary and fourth and fifth maxillary teeth are distinctly fang-like. The lower jaw has fifteen preserved teeth, with three of them being fang-like.[4] By comparison, the teeth in adult Tanystropheus are sharp pegs, while they are tricuspid (bearing three cusps) in juveniles.[5]
Unlike the oval-shaped eye socket of Tanystropheus, the eye socket of Dinocephalosaurus appears to be peach-shaped, with a narrow anterior end. On the top of the skull, the parietal is broad and flattened, bearing no trace of the midline crest found in Tanystropheus.[5] The jugal only has two processes, missing the third backward-projecting process present in most other reptiles.[4] Also missing are the retroarticular process of the posterior (rear) lower jaw (another point of distinction from Tanystropheus),[5] as well as additional teeth and a cavity between the pterygoid bones on the palate.[4]
Neck and trunk
Tanystropheus and Dinocephalosaurus accomplished their extremely elongated necks in different ways. The neck of Tanystropheus is composed of 13 elongated cervical vertebrae,[2] whereas the neck of Dinocephalosaurus is composed of at least 27 cervical vertebrae that are not as elongated. Among the 27 vertebrae of Dinocephalosaurus, the longest is the nineteenth, which measures approximately 91 mm (3.6 in) long. By comparison, the longest vertebrae in a Chinese specimen of Tanystropheus are the ninth and tenth, which measure 248 mm (9.8 in) long. Additionally, the cervical vertebrae of Dinocephalosaurus are not hollow, unlike those of Tanystropheus.[6]
Additional features of the cervical vertebrae which distinguish Dinocephalosaurus from other protorosaurs include the low and keel-like neural spines, and the anterior and posterior (rear) articular surfaces of the vertebrae both being concave (amphicoelous). In the first ten vertebrae, the bottom margin is also concave. The long, slender cervical ribs bear frontal projections free of the vertebral bodies, which are also unique to Dinocephalosaurus. These ribs are aligned along the neck and bridge multiple consecutive vertebral joints, from two or three consecutive joints in the anterior the neck to five or six in the posterior neck. There appear to be no distinct processes on the vertebrae for articulation with the ribs.[4]
The trunk of Dinocephalosaurus has at least 26 vertebrae. In Dinocephalosaurus, there appear to be no lumbar vertebrae, or vertebrae of the trunk lacking ribs. The ribs of the sacrum and tail also do not appear to be fused to their corresponding vertebrae. Each of the gastralia in Dinocephalosaurus is composed of three elements instead of four as in Tanystropheus; they differ in that Dinocephalosaurus only has one element on the midline, while Tanystropheus has two elements that combine to form a midline bar.[4]
Limbs

Dinocephalosaurus had relatively large legs terminating in flipper-like feet. The forelimbs and hindlimbs are roughly the same length, unlike Tanystropheus where the forelimbs are much smaller.[7] Whereas most protorosaurs, such as Tanystropheus, Macrocnemus, and Langobardisaurus, had relatively ossified limbs adapted for terrestrial life, the stout limbs of Dinocephalosaurus are poorly ossified and resemble those of nothosaurs. Out of the carpal bones, only six are ossified; similarly, only three of the tarsal bones are ossified. Additionally, the astragalus and calcaneum also do not articulate with each other in the ankle, instead forming simple and rounded ossifications.[1][4]
These traits are probably neotenic, which similarly characterizes traits found in many other aquatic tetrapods. While Tanystropheus is likely also neotenic, it does not approach Dinocephalosaurus in the extremity of this condition.[4] Several other peculiar traits are present in the feet of Dinocephalosaurus. Unlike Tanystropheus and most other protorosaurs, the fifth metatarsal of Dinocephalosaurus is simple and straight instead of hooked.[1] Tanystropheus merely has a fifth metatarsal which is mildly thickened at the top end, and it additionally possesses a long phalanx on the fifth digit that acts not unlike another metatarsal.[8] On the third digit of the foot, there are four phalanges but none of them appear to be the terminal claw, which suggests that Dinocephalosaurus had a higher-than-average count of at least five phalanges in the third digit.[4]
Discovery and naming
Panxian
.png.webp)
The type specimen of Dinocephalosaurus was first discovered in 2002, during fieldwork conducted in Yangjuan Village, Xinmin District, Panxian County, Guizhou, China. It consists of a nearly-complete skull missing the left side of the jaw, as well as several associated cervical vertebrae. It was subsequently stored at the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP) in Beijing, China under the collection number IVPP V13767, and a research paper describing the specimen was authored by IVPP paleontologist Chun Li and published by Acta Geologica Sinica in December 2003.[5]
A second specimen discovered at the same locality represents a partially articulated skeleton that is only lacking the tail. Likewise stored at the IVPP, the specimen has the collection number IVPP V13898. The specimen was described in a brief correspondence authored by Li, Olivier Rieppel, and Michael LaBarbera that was published by Science in September 2004;[1] a more detailed description was subsequently published by Rieppel, Li, Nicholas Fraser in a 2008 Journal of Vertebrate Paleontology paper.[4]
The Panxian locality, from where these specimens originated, is part of Member II of the Anisian (Middle Triassic) Guanling Formation,[9] which was initially thought to be 230 million years old,[5][10] but has most recently been dated to 244 ± 1.3 million years old based on uranium-lead dating.[11] Predominant deposits at this locality are composed of grey to dark grey marly limestone, as well as cherty limestone containing dolomite and bentonite beds. Dinocephalosaurus was specifically found in layer 90 of the Panxian deposits, a thin limestone layer which is traditionally assigned to the Upper Reptile Horizon (layers 87-90). Further below are the Middle Reptile (layers 81-85) and Lower Reptile (77-79) Horizons.[9]
Luoping
Subsequently, additional Dinocephalosaurus specimens were discovered from the slightly older Luoping locality, which has been dated to 245–244 million years old based on conodont biostratigraphy[12] as well as preliminary radiometric dating.[13] The specimens originate from bed 74 of the Luoping locality, in deposits located near the village of Dawazi, Luoping County, Yunnan, China. Bed 74 is part of a section composed of thin dark grey micritic limestone layers mixed with thicker layers of siliceous limestone and silty limestone, which extends downwards to bed 67.[9][13]
One particular specimen is notable for containing an embryo in its abdominal region, of which cervical vertebrae, forelimbs, and several other elements are preserved. It was collected in 2008; by the time of its collection, weathering had already split the specimen into three blocks, with the gaps having been filled by modern soil. The specimen was then transferred to the Chengdu Center of the China Geological Survey, where it was prepared and stored under the collection number LPV 30280. Subsequently, a description of the specimen, authored by Jun Liu, Chris Organ, Michael Benton, Matthew Brandley, and Jonathan Aitchison, was published in February 2017 by Nature Communications.[13]
Naming
In his 2003 description, Li combined the roots of din- ("terrible"), cephalo- ("head"), and saur ("lizard") to create the genus name Dinocephalosaurus. According to Li, this name refers to the "ghastful skull" of the holotype. Meanwhile, the species name is derived from Latin orientalis ("eastern"), in reference to D. orientalis representing the only known record at the time of the Tanystropheidae in what would have been the eastern portion of the Tethys Ocean.[5]
Classification
Dinocephalosaurus is considered to be a member of the Protorosauria, a group of ubiquitous and diverse Permo-Triassic reptiles.[4] This assignment is based upon characteristics including the low and narrow skull with a short and narrow postorbital region; the long nasal relative to the frontal; the reduced backward projection of the jugal; the presence of more than seven cervical vertebrae, with centra longer than those of the dorsal vertebrae; the low neural spines of the cervical vertebrae; the long cervical ribs; the lack of intercentra articulating with the dorsal centra; and the absence of an entepicondylar foramen on the humerus.[14][15][16][17] These characteristics are shared with Tanystropheus, Macrocnemus, and other protorosaurs.[4]

Protorosaurs were formerly considered to be the ancestors of lizards,[18] but phylogenetic analysis has subsequently verified that they are in fact non-archosaur archosauromorphs.[19] As originally defined, the Protorosauria referred to the group containing Protorosaurus and Prolacerta, and was synonymous with the "Prolacertiformes".[20] In 1997, Nour-Eddine Jalil conducted an analysis of the "Prolacertiformes"; this analysis expanded its definition to include 14 genera, including the Tanystropheidae.[14] Subsequent research has generally found that Prolacerta is closer to the Archosauriformes than Protorosaurus,[21][22] making the "Prolacertiformes" non-monophyletic.[19][23] Whether the remaining protorosaurs – including Protorosaurus, tanystropheids, and drepanosaurs in some studies – form a monophyletic group is unclear. Different phylogenies have supported both monophyletic and non-monophyletic Protorosauria, with the latter arrangement generally having Protorosaurus as being more basal than the Tanystropheidae.[19]
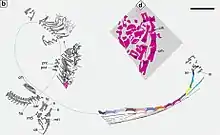
In the original description of Dinocephalosaurus, Li recognized the postorbital region and the elongated cervical centra as being indicative of a close relationship with Tanystropheus, which led him to assign it to the Tanystropheidae.[5] However, Rieppel and colleagues subsequently noted characteristics that distinguished Dinocephalosaurus from derived protorosaurs such as Tanystropheus, Macrocnemus, Langobardisaurus, and Tanytrachelos; these include the lack of tapering at the front end of the nasal and an unreduced contribution of the ilium to the acetabulum.[14] This led Rieppel and colleagues to consider Dinocephalosaurus as an indeterminate protorosaur, with its neck elongation been convergent upon that of Tanystropheus. Their phylogenetic tree, based on a dataset derived from the separate analyses of Jalil (1997),[14] David Dilkes (1998),[17] and Michael Benton & Jackie Allen (1997),[15] is reproduced below, at left.[4]
|
Topology A: Rieppel et al. (2008)[4]
|
Topology B: Liu et al. (2017)[13]
|
|
Liu and colleagues conducted a separate phylogenetic analysis in 2017. They criticized the analysis of Rieppel and colleagues as having unnecessarily repeated several characters in their data, thus imbuing the repeated characters with undue weight in the analysis. In their own analysis, Liu and colleagues used the same source datasets, but deleted repeated characters, added two new characters from an analysis by Sean Modesto and Hans-Dieter Sues (2004),[21] and removed poorly preserved or potentially chimeric taxa such as Cosesaurus, Kadimakara, Trachelosaurus, and Malerisaurus. Analyses based on parsimony-based and Bayesian methods found that Dinocephalosaurus was a member of the Tanystropheidae, being the sister group to a clade containing Tanystropheus, Macrocnemus, Langobardisaurus, and Tanytrachelos. The analyses also recovered a monophyletic Protorosauria, albeit to the exclusion of Prolacerta. The tree from the parsimony-based analysis is reproduced above, at right.[13]
Paleobiology
Neck and feeding

The long neck of Dinocephalosaurus probably served a functional role. In particular, the length of the neck places a long distance between the head and the remainder of the body. This would have allowed Dinocephalosaurus to approach potential prey without the majority of its bulk being detected, which would have been effective in the murky waters of its habitat.[1]
Dinocephalosaurus may also have used its neck to capture its prey via suction feeding. After flexing its neck to the side (which would have been facilitated by the slenderness of the cervical ribs), the act of straightening the neck would have caused the cervical ribs to splay outwards due to the action of the neck muscles attached to the ribs. As the head lunged forward, the volume of the esophagus would have increased, creating suction. Once the prey was caught, the fang-like teeth would have secured the prey in the mouth.[1] Like other aquatic amniotes, Dinocephalosaurus would have swallowed and digested its prey head-first, as evidenced by the preservation of a perleidid fish in the abdominal region of LPV 30280 from Luoping.[13][24]
It is not likely that Dinocephalosaurus used its long neck for breathing by extending it vertically. If it tried to do so, the difference in pressure between the surface and its torso would be sufficiently extreme such that its lungs would not have inflated. Thus, in order to breathe, Dinocephalosaurus would need to approach the surface with a nearly horizontal neck. The horizontal posture of the neck would also have facilitated locomotion at the surface, due to its long profile increasing its "hull length" and reducing the effect of resistance from waves.[1]
Reproduction
Dinocephalosaurus represents the climax of aquatic adaptations among the protorosaurs.[1] Given its long neck and paddle-like limbs, it was probably incapable of functioning comfortably in a terrestrial environment. One consequence of these adaptations is that Dinocephalosaurus would have been incapable of building nests on land. This would also have prevented it from possessing hard-shelled reptilian eggs; such eggs necessitate the exchange of gases with the outside environment, and this process is significantly slower in water than it is in air.[25] Thus, Dinocephalosaurus could not have been oviparous. At the same time, however, archosaurs are known for the total absence of viviparous, or live birth, among its living members.[13][26]

The embryonic individual of Dinocephalosaurus preserved inside LPV 30280 from Luoping can be identified as such for several reasons. First, it is enclosed entirely within the body cavity of the adult. Its cervical ribs - which are long, like that of the adult - face the same direction as the dorsal vertebrae of the adult, which is in contradiction to the typical head-first method of swallowing prey among amniotes. It is also preserved with its neck curling towards its forelimbs, a posture which is seen among vertebrate embryos but not adult Dinocephalosaurus specimens, which generally have the neck bent upwards.[4] The absence of hands associated with the forelimbs in the embryo may be an artifact of preservation, but it may also be due to the sequence in which the limb bones of tetrapods ossify.[13]
Although the presence of an embryo per se would be equivocal regarding this issue, the conditions in which the embryo were preserved provide strong evidence that Dinocephalosaurus was viviparous. There is no preserved calcified eggshell surrounding the embryo, despite the presence of delicately-preserved calcified elements from other animals at Luoping.[9] This is consistent with the eggshells surrounding the embryos of viviparous reptiles being non-calcified membranes.[27] Furthermore, the relative proportions of the humerus and the fibula in the embryo, compared with the maternal individual and IVPP V13898 from Panxian, indicate that the embryo is around 12% of the size of its mother. Combined with its ossified bones, this suggests that the embryo was at an advanced developmental stage, whereas crocodilians, birds, turtles, and tuataras lay eggs at very early developmental stages.[13][28]
Overall, the evidence provided by the embryo suggests that Dinocephalosaurus was viviparous, making it the first viviparous archosauromorph[13] asides from possibly metriorhynchids.[29] This is consistent with the separation of its sacral ribs from its sacrum,[4] which indicates a mobile pelvis that could have functioned in giving birth.[30][31] While the otherwise absence of viviparous archosaurs has been historically attributed to common attributes inherited from the archosaurian stem-lineage, the discovery that Dinocephalosaurus was viviparous suggests that this phenomenon is due to lineage-specific lifestyle restrictions. Although the sex-determination systems among living archosauromorphs are diverse, with crocodilians and turtles using temperature-dependent sex determination,[32] phylogenetic modelling suggests that Dinocephalosaurus retained the basal condition of genotypic sex determination from early diapsids, and that this system facilitated its transition to an obligately marine lifestyle alongside viviparity.[13][33]
Paleoecology
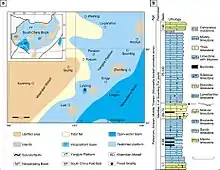
Until at least the end of the Middle Triassic, high sea levels enabled shallow water to cover much of the South China Block,[34][35] a tectonic plate that today consists of the stable Yangtze Craton and the less stable South China Fold Belt. A mountain-building event known as the Indosinian orogeny uplifted Precambrian rocks to form four major landmasses on the South China Block: Khamdian to the west, Jiangnan in the centre, Yunkai to the south, and Cathaysia to the east. Island chains also stretched between Yunkai and Cathaysia in the east. Located between Khamdian and Jiangnan was a deep oceanic basin known as the Nanpanjiang Basin.[35][36][37] Along the western edge of this basin, fossil-bearing sediments were laid down to become what are now the Lagerstätten (sedimentary deposits characterized by exemplary preservation) of Panxian, Luoping, and other localities.[9]

At Panxian, there appears to have been a transition in the endemic marine reptile fauna, possibly caused by volcanism. This volcanism is indicated by the presence of a bentonite layer between the Middle and Upper Reptile Horizons.[38] From lower in the Upper Reptile Horizon, fossils have also been found of the ubiquitous mixosaurian ichthyosaur Mixosaurus panxianensis (which occurs in all layers), the pachypleurosaurs Keichousaurus sp. and Wumengosaurus delicatomandibularis, and fish, although fossils of the latter are fragmentary. The lower Middle and Lower Reptile Horizons also include the mixosaurian Phalarodon cf. fraasi, the primitive ichthyosaur Xinminosaurus catactes, the placodont Placodus inexpectatus, the nothosaurs Lariosaurus hongguoensis and Nothosaurus yangjuanensis, and the archosaur Qianosuchus mixtus, alongside bivalves and saurichthyid fish.[9][39]
The fauna of Luoping appears to have been preserved in a small intraplatform basin instead of the surrounding open water, judging by the anoxic sediments present at the site.[9] Out of 19,759 specimens, 93.7% of the fossils found at Luoping are arthropods: decapods, isopods, crab-like cycloids, shrimp-like mysidaceans, clam shrimp, and ostracods, as well as rare millipedes and horseshoe crabs. By comparison, only 0.07% of specimens come from marine reptiles,[40] which include Dinocephalosaurus alongside Mixosaurus cf. panxianensis, Dianopachysaurus dingi, Sinosaurosphargis yunguiensis, and an archosaur.[9] Otherwise, fish including saurichthyids, palaeoniscids, birgeriids, perleidids, eugnathids, semionotids, pholidopleurids, peltopleurids, and coelacanths have been found at Luoping, forming 3.66% of fossils with 25 taxa in 9 families. Molluscs such as bivalves and gastropods, along with ammonoids and belemnoids, account for 1.69% of specimens. Rare and probably non-endemic fossils also include echinoderms such as crinoids, starfish, and sea urchins; branchiopods; and relatively complete conifer branches and leaves, which probably originated from coastal forests less than 10 km (6.2 mi) away.[40]
References
- Li, C.; Rieppel, O.; LaBarbera, M.C. (2004). "A Triassic Aquatic Protorosaur with an Extremely Long Neck". Science. 305 (5692): 1931. doi:10.1126/science.1100498. PMID 15448262. S2CID 38739295.
- Rieppel, O.; Jiang, D.-Y.; Fraser, N.C.; Hao, W.-C.; Motani, R.; Sun, Y.-L.; Sun, Z.-Y. (2010). "Tanystropheus cf. T. longobardicus from the Early Late Triassic of Guizhou Province, Southwestern China". Journal of Vertebrate Paleontology. 30 (4): 1082–1089. doi:10.1080/02724634.2010.483548. S2CID 86315078.
- Cloudsley-Thompson, J.L. (2005). "Therapsids, Anapsids and Early Diapsids". Ecology and Behaviour of Mesozoic Reptiles. Berlin: Springer-Verlag. pp. 113–123. doi:10.1007/3-540-26571-6_8. ISBN 978-3-540-26571-9.
- Rieppel, O.; Li, C.; Fraser, N.C. (2008). "The Skeletal Anatomy of the Triassic Protorosaur Dinocephalosaurus orientalis Li, from the Middle Triassic of Guizhou Province, Southern China". Journal of Vertebrate Paleontology. 28 (1): 95–110. doi:10.1671/0272-4634(2008)28[95:tsaott]2.0.co;2. JSTOR 30126337.
- Li, C. (2003). "First record of protorosaurid reptile (Order Protorosauria) from the Middle Triassic of China". Acta Geologica Sinica - English Edition. 77 (4): 419–423. doi:10.1111/j.1755-6724.2003.tb00122.x.
- Rieppel, O. (2001). "A new species of Tanystropheus (Reptilia: Protorosauria) from the Middle Triassic of Makhtesh Ramon, Israel". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 221 (2): 271–287. doi:10.1127/njgpa/221/2001/271.
- Renesto, S. (2005). "A new specimen of Tanystropheus (Reptilia Protorosauria) from the Middle Triassic of Switzerland and the ecology of the genus". Rivista Italiana di Paleontologia e Stratigrafia. 111 (3): 377–394. doi:10.13130/2039-4942/6327.
- Renesto, S. (1994). "A new prolacertiform reptile from the Late Triassic of Northern Italy". Rivista Italiana di Paleontologia e Stratigrafia. 100 (2): 285–306. doi:10.13130/2039-4942/8615.
- Benton, M.J.; Zhang, Q.; Hu, S.; Chen, Z.-Q.; Wen, W.; Liu, J.; Zhou, C.; Xie, T.; Tong, J.; Choo, B. (2013). "Exceptional vertebrate biotas from the Triassic of China, and the expansion of marine ecosystems after the Permo-Triassic mass extinction". Earth-Science Reviews. 123: 199–243. Bibcode:2013ESRv..125..199B. doi:10.1016/j.earscirev.2013.05.014.
- Sun, Z.; Sun, Y.; Hao, W.; Jiang, D. (2006). "Conodont Evidence for the Age of the Panxian Fauna, Guizhou, China". Acta Geologica Sinica - English Edition. 80 (5): 621–630. doi:10.1111/j.1755-6724.2006.tb00284.x.
- Wang, Y.; Yang, D.; Han, J.; Wang, L.; Yao, J.; Liu, D. (2014). "The Triassic U–Pb age for the aquatic long-necked protorosaur of Guizhou, China". Geological Magazine. 151 (4): 749–754. Bibcode:2014GeoM..151..749W. doi:10.1017/S001675681400003X.
- Zhang, Q.-Y.; Zhou, C.-Y.; Lu, T.; Xie, T.; Lou, X.-Y.; Liu, W.; Sun, Y.-Y.; Huang, J.-Y.; Zhao, L.-S. (2009). "A conodont-based Middle Triassic age assignment for the Luoping Biota of Yunnan, China". Science in China Series D: Earth Sciences. 52 (10): 1673–1678. Bibcode:2009ScChD..52.1673Z. doi:10.1007/s11430-009-0114-z. S2CID 129857181.
- Liu, J.; Organ, C.L.; Benton, M.J.; Brandley, M.C.; Aitchison, J.C. (2017). "Live birth in an archosauromorph reptile". Nature Communications. 8: 14445. Bibcode:2017NatCo...814445L. doi:10.1038/ncomms14445. ISSN 2041-1723. PMC 5316873. PMID 28195584.
- Jalil, N.-E. (1997). "A new prolacertiform diapsid from the Triassic of North Africa and the interrelationships of the Prolacertiformes". Journal of Vertebrate Paleontology. 17 (3): 506–525. doi:10.1080/02724634.1997.10010998.
- Benton, M.J.; Allen, J.L. (1997). "Boreopricea from the Lower Triassic of Russia, and the relationships of the prolacertiform reptiles". Palaeontology. 40: 931–953.
- Evans, S.E. (1998). "The early history and relationships of the Diapsida". In Benton, M.J. (ed.). The Phylogeny and Classification of the Tetrapods. 1. Oxford: Clarendon Press. pp. 221–253.
- Dilkes, D.W. (1998). "The early Triassic rhynchosaur Mesosuchus browni and the interrelationships of basal archosauromorph reptiles". Philosophical Transactions of the Royal Society B. 353 (1368): 501–541. doi:10.1098/rstb.1998.0225. PMC 1692244.
- Parrington, F.R. (1935). "On Prolacerta broomi, gen. et sp. n., and the origin of lizards". Annals and Magazine of Natural History. 10. 16 (92): 197–205. doi:10.1080/00222933508655037.
- Ezcurra, M.D. (2016). "The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms". PeerJ. 4: e1778. doi:10.7717/peerj.1778. PMC 4860341. PMID 27162705.
- Camp, C.L. (1945). "Prolacerta and the protorosaurian reptiles; Part I". American Journal of Science. 243 (1): 17–32. Bibcode:1945AmJS..243...17C. doi:10.2475/ajs.243.1.17.
- Modesto, S.P.; Sues, H.D. (2004). "The skull of the Early Triassic archosauromorph reptile Prolacerta broomi and its phylogenetic significance". Zoological Journal of the Linnean Society. 140 (3): 335–351. doi:10.1111/j.1096-3642.2003.00102.x.
- Gottmann-Quesada, A.; Sander, P.M. (2009). "A redescription of the early archosauromorph Protorosaurus speneri Meyer, 1832, and its phylogenetic relationships". Palaeontographica Abteilung A. 287 (4–6): 123–200. doi:10.1127/pala/287/2009/123.
- Ezcurra, M.D.; Scheyer, T.M.; Butler, R.J. (2014). "The origin and early evolution of Sauria: reassessing the Permian saurian fossil record and the timing of the crocodile-lizard divergence". PLOS ONE. 9 (2): e89165. Bibcode:2014PLoSO...989165E. doi:10.1371/journal.pone.0089165. PMC 3937355. PMID 24586565.
- Caldwell, M.W.; Lee, M.S.Y. (2001). "Live birth in Cretaceous marine lizards (mosasauroids)". Proceedings of the Royal Society B. 268 (1484): 2397–2401. doi:10.1098/rspb.2001.1796. PMC 1088892. PMID 11747556.
- Shine, R. (1985). "The Evolution of Viviparity in Reptiles: An Ecological Analysis". In Gans, C.; Billett, F. (eds.). Biology of the Reptilia. 15. New York: John Wiley & Sons. pp. 605–694.
- Blackburn, D.G.; Evans, H.E. (1986). "Why are there no viviparous birds?". The American Naturalist. 128 (2): 165–190. doi:10.1086/284552. JSTOR 2461543. S2CID 84158031.
- Stewart, J.R.; Thompson, M.B. (2009). "Parallel Evolution of Placentation in Australian Scincid Lizards". Journal of Experimental Zoology Part B: Molecular and Developmental Evolution. 312B (6): 590–602. doi:10.1002/jez.b.21245. PMID 19012323.
- Andrews, R.M. (2004). "Embryonic development". In Deeming, D.C. (ed.). Reptilian Incubation: Environment, Evolution, and Behaviour. Nottingham: Nottingham University Press. pp. 75–102.
- Herrera, Y.; Fernandez, M.S.; Lamas, S.G.; Campos, L.; Talevi, M.; Gasparini, Z. (2017). "Morphology of the sacral region and reproductive strategies of Metriorhynchidae: a counter-inductive approach". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 106 (4): 247–255. doi:10.1017/S1755691016000165.
- Cheng, Y.-N.; Wu, X.-C.; Ji, Q. (2004). "Triassic marine reptiles gave birth to live young". Nature. 432 (7015): 383–386. Bibcode:2004Natur.432..383C. doi:10.1038/nature03050. PMID 15549103. S2CID 4391810.
- O'Keefe, F.R.; Chiappe, L.M. (2011). "Viviparity and K-Selected Life History in a Mesozoic Marine Plesiosaur (Reptilia, Sauropterygia)". Science. 333 (6044): 870–873. Bibcode:2011Sci...333..870O. doi:10.1126/science.1205689. PMID 21836013. S2CID 36165835.
- Organ, C.L.; Janes, D.E. (2008). "Evolution of sex chromosomes in Sauropsida". Integrative and Comparative Biology. 48 (4): 512–519. doi:10.1093/icb/icn041. PMC 4553705. PMID 21669812.
- Organ, C.L.; Janes, D.E.; Meade, A.; Pagel, M. (2009). "Genotypic sex determination enabled adaptive radiations of extinct marine reptiles". Nature. 461 (7262): 389–392. Bibcode:2009Natur.461..389O. doi:10.1038/nature08350. PMID 19759619. S2CID 351047.
- Lehrmann, D.J.; Enos, P.; Payne, J.L.; Montgomery, P.; Wei, J.; Yu, Y.; Xiao, J.; Orchard, M.J. (2005). "Permian and Triassic depositional history of the Yangtze platform and Great Bank of Guizhou in the Nanpanjiang basin of Guizhou and Guangxi, South China" (PDF). Albertiana. 33: 149–168.
- Lehrmann, D.J.; Minzoni, M.; Enos, P.; Yu, Y.-Y.; Wei, J.-Y.; Li, R.-X. (2009). "Triassic depositional history of the Yangtze platform and Great Bank of Guizhou in the Nanpanjiang basin of South China". Journal of Earth Sciences and Environment. 31 (4): 344–367.
- Lehrmann, D.J.; Payne, J.L.; Felix, S.V.; Dillett, P.M.; Wang, H.; Yu, Y.; Wei, J. (2003). "Permian–Triassic Boundary Sections from Shallow-Marine Carbonate Platforms of the Nanpanjiang Basin, South China: Implications for Oceanic Conditions Associated with the End-Permian Extinction and Its Aftermath". PALAIOS. 18 (2): 138–152. Bibcode:2003Palai..18..138L. CiteSeerX 10.1.1.486.1486. doi:10.1669/0883-1351(2003)18<138:PBSFSC>2.0.CO;2.
- Lehrmann, D.J.; Payne, J.L.; Pei, D.; Enos, P.; Druke, D.; Steffen, K.; Zhang, J.; Wei, J.; Orchard, M.J.; Ellwood, B. (2007). "Record of the end-Permian extinction and Triassic biotic recovery in the Chongzuo-Pingguo platform, southern Nanpanjiang basin, Guangxi, South China". Palaeogeography, Palaeoclimatology, Palaeoecology. 252 (1): 200–217. Bibcode:2007PPP...252..200L. doi:10.1016/j.palaeo.2006.11.044.
- Motani, R.; Jiang, D.-Y.; Tintori, A.; Sun, Y.-L.; Hao, W.-C.; Boyd, A.; Sanja, H.-F.; Schmitz, L.; Shin, Y.-J.; Sun, Z.-Y. (2008). "Horizons and Assemblages of Middle Triassic Marine Reptiles from Panxian, Guizhou, China". Journal of Vertebrate Paleontology. 28 (3): 900–903. doi:10.1671/0272-4634(2008)28[900:HAAOMT]2.0.CO;2.
- Jiang, D.; Motani, R.; Hao, W.; Rieppel, O.; Sun, Y.; Tintori, A.; Sun, Z.; Schmitz, L. (2009). "Biodiversity and Sequence of the Middle Triassic Panxian Marine Reptile Fauna, Guizhou Province, China". Acta Geologica Sinica - English Edition. 83 (3): 451–459. doi:10.1111/j.1755-6724.2009.00047.x.
- Hu, S.-X.; Zhang, Q.-Y.; Chen, Z.-Q.; Zhou, C.-Y.; Lü, T.; Xie, T.; Wen, W.; Huang, J.-Y.; Benton, M.J. (2008). "The Luoping biota: exceptional preservation, and new evidence on the Triassic recovery from end-Permian mass extinction". Proceedings of the Royal Society B. 278 (1716): 2274–2282. doi:10.1098/rspb.2010.2235. PMC 3119007. PMID 21183583.
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||