TRPA (ion channel)
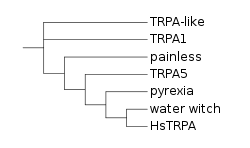
TRPA is a family of transient receptor potential ion channels. The TRPA family is made up of 7 subfamilies: TRPA1, TRPA- or TRPA1-like, TRPA5, painless, pyrexia, waterwitch, and HsTRPA. TRPA1 is the only subfamily widely expressed across animals, while the other subfamilies (collectively referred to as the basal clade) are largely absent in deuterostomes (and in the case of HsTRPA, only expressed in hymenopteran insects).[2][1][3][4]
| transient receptor potential cation channel, subfamily A, member 1 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | TRPA1 | ||||||
| Alt. symbols | ANKTM1 | ||||||
| IUPHAR | 485 | ||||||
| NCBI gene | 8989 | ||||||
| HGNC | 497 | ||||||
| OMIM | 604775 | ||||||
| RefSeq | NM_007332 | ||||||
| UniProt | O75762 | ||||||
| Other data | |||||||
| Locus | Chr. 8 q13 | ||||||
| |||||||
TRPA1s have been the most extensively studied subfamily; they typically contain 14 N-terminal ankyrin repeats and are believed to function as mechanical stress, temperature, and chemical sensors. TRPA1 is known to be activated by compounds such as isothiocyanate (which are the pungent chemicals in substances such as mustard oil and wasabi) and Michael acceptors (e.g. cinnamaldehyde). These compounds are capable of forming covalent chemical bonds with the protein's cysteins.[5] Non-covalent activators of TRPA1 also exists, such as methyl salicylate, menthol, and the synthetic compound PF-4840154.[6][1][7]
The thermal sensitivity of TRPAs varies by species. For example, TRPA1 functions as a high-temperature sensor in insects and snakes, but as a cold sensor in mammals.[8] The basal TRPAs have evolved some degree of thermal sensitivity as well: painless and pyrexia function in high-temperature sensing in Drosophila melanogaster, and the honey bee HsTRPA underwent neofunctionalization following its divergence from waterwitch, gaining function as a high-temperature sensor.[9]
TRPA1s promiscuity with respect to sensory modality has been the source of controversy, particularly when considering its ability to detect cold.[10] More recent work has alternatively (or additionally) proposed that reactive oxygen species activate TRPA1, across species.[11][12]
References
- Himmel NJ, Letcher JM, Sakurai A, Gray TR, Benson MN, Cox DN (November 2019). "Drosophila menthol sensitivity and the Precambrian origins of transient receptor potential-dependent chemosensation". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 374 (1785): 20190369. doi:10.1098/rstb.2019.0369. PMC 6790378. PMID 31544603.
- Kang K, Pulver SR, Panzano VC, Chang EC, Griffith LC, Theobald DL, Garrity PA (March 2010). "Analysis of Drosophila TRPA1 reveals an ancient origin for human chemical nociception". Nature. 464 (7288): 597–600. Bibcode:2010Natur.464..597K. doi:10.1038/nature08848. PMC 2845738. PMID 20237474.
- Peng G, Shi X, Kadowaki T (March 2015). "Evolution of TRP channels inferred by their classification in diverse animal species". Molecular Phylogenetics and Evolution. 84: 145–57. doi:10.1016/j.ympev.2014.06.016. PMID 24981559.
- Kozma MT, Schmidt M, Ngo-Vu H, Sparks SD, Senatore A, Derby CD (2018). "Chemoreceptor proteins in the Caribbean spiny lobster, Panulirus argus: Expression of Ionotropic Receptors, Gustatory Receptors, and TRP channels in two chemosensory organs and brain". PLOS ONE. 13 (9): e0203935. Bibcode:2018PLoSO..1303935K. doi:10.1371/journal.pone.0203935. PMC 6150509. PMID 30240423.
- Nilius B, Owsianik G, Voets T, Peters JA (January 2007). "Transient receptor potential cation channels in disease". Physiological Reviews. 87 (1): 165–217. doi:10.1152/physrev.00021.2006. PMID 17237345.
- Ryckmans T, Aubdool AA, Bodkin JV, Cox P, Brain SD, Dupont T, et al. (August 2011). "Design and pharmacological evaluation of PF-4840154, a non-electrophilic reference agonist of the TrpA1 channel". Bioorganic & Medicinal Chemistry Letters. 21 (16): 4857–9. doi:10.1016/j.bmcl.2011.06.035. PMID 21741838.
- Zygmunt PM, Högestätt ED (2014). "TRPA1". Handbook of Experimental Pharmacology. 222: 583–630. doi:10.1007/978-3-642-54215-2_23. ISBN 978-3-642-54214-5. PMID 24756722.
- Panzano VC, Kang K, Garrity PA (June 2010). "Infrared snake eyes: TRPA1 and the thermal sensitivity of the snake pit organ". Science Signaling. 3 (127): pe22. doi:10.1126/scisignal.3127pe22. PMID 20571127. S2CID 13504270.
- Kohno K, Sokabe T, Tominaga M, Kadowaki T (September 2010). "Honey bee thermal/chemical sensor, AmHsTRPA, reveals neofunctionalization and loss of transient receptor potential channel genes". The Journal of Neuroscience. 30 (37): 12219–29. doi:10.1523/JNEUROSCI.2001-10.2010. PMC 6633439. PMID 20844118.
- Caspani O, Heppenstall PA (March 2009). "TRPA1 and cold transduction: an unresolved issue?". The Journal of General Physiology. 133 (3): 245–9. doi:10.1085/jgp.200810136. PMC 2654088. PMID 19237589.
- Arenas OM, Zaharieva EE, Para A, Vásquez-Doorman C, Petersen CP, Gallio M (December 2017). "Activation of planarian TRPA1 by reactive oxygen species reveals a conserved mechanism for animal nociception". Nature Neuroscience. 20 (12): 1686–1693. doi:10.1038/s41593-017-0005-0. PMC 5856474. PMID 29184198.
- Miyake T, Nakamura S, Zhao M, So K, Inoue K, Numata T, et al. (September 2016). "Cold sensitivity of TRPA1 is unveiled by the prolyl hydroxylation blockade-induced sensitization to ROS". Nature Communications. 7: 12840. Bibcode:2016NatCo...712840M. doi:10.1038/ncomms12840. PMC 5027619. PMID 27628562.
External links
- TRPA1+protein,+human at the US National Library of Medicine Medical Subject Headings (MeSH)
- "TRIP Database". a manually curated database of protein-protein interactions for mammalian TRP channels.