GPR17
Uracil nucleotide/cysteinyl leukotriene receptor is a G protein-coupled receptor that in humans is encoded by the GPR17 gene located on chromosome 2 at position q21.[5][6] The actual activating ligands for and some functions of this receptor are disputed.


History
Initially discovered in 1998 as an Orphan receptor, i.e. a receptor whose activating ligand(s) and function were unknown, GPR17 was "deorphanized" in a study that reported it to be a receptor for LTC4, LTD4, and uracil nucleotides.[7] In consequence, GPR17 attracted attention as a potential mediator of reactions caused by LTC4 and LTD4 viz., asthma, rhinitis, and urticarial triggered by allergens, nonsteroidal anti-inflammatory drugs, and exercise (see Aspirin-induced asthma). Subsequent reports, however, have varied in results: studies focusing on the allergen and non-allergen reactions find that GPR17-bearing cells do not respond to LTC4, LTD4, and uracil nucleotides[8] while studies focusing on nerve tissue find that certain types of GPR17-bearing oligodendrocytes do indeed respond to them.[7] In 2013 and 2014 reports, the International Union of Basic and Clinical Pharmacology took no position on which of these are true ligands for GPR17.[9][10] GPR17 is a constitutively active receptor, i.e. a receptor that has baseline activity which is independent of, although potentially increased by, its ligands.[9]
Biochemistry
GPR17 has a structure which is intermediate between the cysteinyl leukotriene receptor group (i.e. cysteinyl leukotriene receptor 1 and cysteinyl leukotriene receptor 2) and the purine P2Y subfamily of 12 receptors (see P2Y receptors), sharing 28 to 48% amino acid identity with them. GPR17 is a G protein coupled receptor that acts primarily through G proteins linked to the Gi alpha subunit but also to Gq alpha subunit.[7][11] Matching these structural relationships, GPR17 has been reported to be activated by cysteinyl leukotrienes (i.e. LTC4 and LTD4) as well as the purines (i.e., uridine, Uridine diphosphate (UDP), UDP-glucose). Further relating these receptors, GPR17 may dimerize (i.e. associate with) certain of the cited cysteinyl leukotriene or purine receptors in mediating cell responses and this dimerization may explain some of the discrepancies reported for the ability of these ligands to activate GPR17 as expressed in different cell types (see below section of Function). GPR17 is also activated by the emergency-signaling and atherosclerosis-promoting oxysterols and by synthetic compounds with broadly different structures. Relevant to its activating ligands as well as its reported interaction with other G protein coupled receptors, GPR17 is a promiscuous receptor.[7]
Montelukast which inhibits cysteinyl leukotriene receptor 1 and is in clinical use for the chronic and preventative treatment of LTC4- and LTD4-promoted allergic and non-allergic diseases, and Cangrelor, which inhibits P2Y purinergic receptors and is approved in the USA as an antiplatelet drug, inhibit the GPR17 receptor.[7]
Distribution
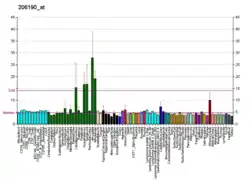
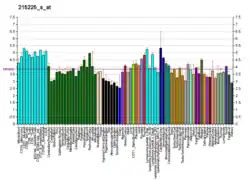
GPR17 was first clone form and is highly expressed in certain precursors of oligodendrocytes in the nerve tissue of the central nervous system (CNS); it is overexpress in CNS tissues experiencing demyelination injuries; within 48 hours of the latter types of injuries, GPR17 expression is induced in dying neurons within and on the borders of injury, in infiltrating microglia and macrophages, and in activated oligodendrocyte precursor cells.[7]
Function
Studies focusing on allergic and hypersensitivity reactions have found that the LTC4 and LTD4 ligands for Cysteinyl leukotriene receptor 1 (CysLTR1) and Cysteinyl leukotriene receptor 2, which mediate these reactions, have disputed findings that LTC4 and LTD4 are ligands for GPR17. They have shown that cells co-expressing both CysLTR1 and GPR17 receptors exhibit a marked reduction in binding LTC4 and that mice lacking GPR17 are hyper-responsive to igE-induced passive cutaneous anaphylaxis. They therefore have nominated GPR17 as functioning to inhibit CysLTR1 in these model systems and as such might serve to dampen the acute reactions involving the cited LTs.[12]
Studies focusing on nerve tissue indicate that GPR17 is: a) highly expressed in precursors to mature oligodendrocytes but not expressed in mature oligodendrocytes, suggesting that GPR17 must be down-regulated in order for precursor cells to proceed to terminal oligodendrocyte differentiation; b) activated by uridine, Uridine diphosphate (UDP) and UDP-glucose to stimulate outward K+ channels and the aforementioned maturation responses in oligodenrocyte precursor cells; c) also activated by LTC4 and LTD4; d) more highly expressed in central nervous system (CNS) tissues of animal models undergoing ischemia, Experimental autoimmune encephalomyelitis, and focal demyelination as well as in the CNS tissues of humans suffering brain damage due to ischemia, trauma, and multiple sclerosis; e) expressed in injured neurons and associated with the rapid death and clearance of these neurons in a model of mouse spinal cord crush injury; f) acts to reduce the extent of spinal cord injury in the latter model based on the increased extent of injury in GPR17-depleted mice; and g) acts to reduce inflammation, elevate hippocampus neurogenesis, and improve learning and memory in a rat model of age-related cognitive impairment based on the effects of the GPR17 antagonist, montelukast, as well as of GPR17 depletion. The studies suggest that GPR17 is a sensor of damage in the CNS and participates in the resolution of this damage by clearing and/or promoting the re-myelination of injured neurons caused by a variety of insults perhaps including old age.[7][13][14][15]
The GPR17 gene has also been found to regulate food intake response mediated by FOXO1.[16]
Clinical significance
GPR17 has been proposed as a potential pharmacological target for the treatment of multiple sclerosis and traumatic brain injury in humans.[7][15][17]
References
- GRCh38: Ensembl release 89: ENSG00000144230 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000052229 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Raport CJ, Schweickart VL, Chantry D, Eddy RL, Shows TB, Godiska R, Gray PW (January 1996). "New members of the chemokine receptor gene family". Journal of Leukocyte Biology. 59 (1): 18–23. doi:10.1002/jlb.59.1.18. PMID 8558062. S2CID 24239900.
- "Entrez Gene: GPR17 G protein-coupled receptor 17".
- Fumagalli M, Lecca D, Abbracchio MP (2016). "CNS remyelination as a novel reparative approach to neurodegenerative diseases: The roles of purinergic signaling and the P2Y-like receptor GPR17". Neuropharmacology. 104: 82–93. doi:10.1016/j.neuropharm.2015.10.005. PMID 26453964. S2CID 26235050.
- Kanaoka Y, Boyce JA (2014). "Cysteinyl leukotrienes and their receptors; emerging concepts". Allergy, Asthma & Immunology Research. 6 (4): 288–95. doi:10.4168/aair.2014.6.4.288. PMC 4077954. PMID 24991451.
- Davenport AP, Alexander SP, Sharman JL, Pawson AJ, Benson HE, Monaghan AE, Liew WC, Mpamhanga CP, Bonner TI, Neubig RR, Pin JP, Spedding M, Harmar AJ (2013). "International Union of Basic and Clinical Pharmacology. LXXXVIII. G protein-coupled receptor list: recommendations for new pairings with cognate ligands". Pharmacological Reviews. 65 (3): 967–86. doi:10.1124/pr.112.007179. PMC 3698937. PMID 23686350.
- Bäck M, Powell WS, Dahlén SE, Drazen JM, Evans JF, Serhan CN, Shimizu T, Yokomizo T, Rovati GE (2014). "Update on leukotriene, lipoxin and oxoeicosanoid receptors: IUPHAR Review 7". British Journal of Pharmacology. 171 (15): 3551–74. doi:10.1111/bph.12665. PMC 4128057. PMID 24588652.
- Wódz-Naskiewicz K, Pawliczak R (2009). "[Cysteinyl leukotrienes and their receptors]". Postepy Biochemii (in Polish). 55 (4): 395–405. PMID 20201353.
- Brink C, Dahlén SE, Drazen J, Evans JF, Hay DW, Rovati GE, Serhan CN, Shimizu T, Yokomizo T (March 2004). "International Union of Pharmacology XLIV. Nomenclature for the oxoeicosanoid receptor". Pharmacological Reviews. 56 (1): 149–57. doi:10.1124/pr.56.1.4. PMID 15001665. S2CID 7229884.W
- Ceruti S, Villa G, Genovese T, Mazzon E, Longhi R, Rosa P, Bramanti P, Cuzzocrea S, Abbracchio MP (2009). "The P2Y-like receptor GPR17 as a sensor of damage and a new potential target in spinal cord injury". Brain. 132 (Pt 8): 2206–18. doi:10.1093/brain/awp147. PMID 19528093.
- Marschallinger J, Schäffner I, Klein B, Gelfert R, Rivera FJ, Illes S, Grassner L, Janssen M, Rotheneichner P, Schmuckermair C, Coras R, Boccazzi M, Chishty M, Lagler FB, Renic M, Bauer HC, Singewald N, Blümcke I, Bogdahn U, Couillard-Despres S, Lie DC, Abbracchio MP, Aigner L (2015). "Structural and functional rejuvenation of the aged brain by an approved anti-asthmatic drug". Nature Communications. 6: 8466. doi:10.1038/ncomms9466. PMC 4639806. PMID 26506265.
- Burnstock G (May 2016). "An introduction to the roles of purinergic signalling in neurodegeneration, neuroprotection and neuroregeneration". Neuropharmacology. 104: 4–17. doi:10.1016/j.neuropharm.2015.05.031. PMID 26056033. S2CID 38914606.
- Ren H, Orozco IJ, Su Y, Suyama S, Gutiérrez-Juárez R, Horvath TL, Wardlaw SL, Plum L, Arancio O, Accili D (June 2012). "FoxO1 target Gpr17 activates AgRP neurons to regulate food intake". Cell. 149 (6): 1314–26. doi:10.1016/j.cell.2012.04.032. PMC 3613436. PMID 22682251.
- Chen Y, Wu H, Wang S, Koito H, Li J, Ye F, Hoang J, Escobar SS, Gow A, Arnett HA, Trapp BD, Karandikar NJ, Hsieh J, Lu QR (2009). "The oligodendrocyte-specific G protein-coupled receptor GPR17 is a cell-intrinsic timer of myelination". Nature Neuroscience. 12 (11): 1398–406. doi:10.1038/nn.2410. PMC 2783566. PMID 19838178.
Further reading
- Macrez-Leprêtre N, Kalkbrenner F, Morel JL, Schultz G, Mironneau J (April 1997). "G protein heterotrimer Galpha13beta1gamma3 couples the angiotensin AT1A receptor to increases in cytoplasmic Ca2+ in rat portal vein myocytes". The Journal of Biological Chemistry. 272 (15): 10095–102. doi:10.1074/jbc.272.15.10095. PMID 9092554.
- Bläsius R, Weber RG, Lichter P, Ogilvie A (April 1998). "A novel orphan G protein-coupled receptor primarily expressed in the brain is localized on human chromosomal band 2q21". Journal of Neurochemistry. 70 (4): 1357–65. doi:10.1046/j.1471-4159.1998.70041357.x. PMID 9523551. S2CID 10174931.
- Rual JF, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N, Berriz GF, Gibbons FD, Dreze M, Ayivi-Guedehoussou N, Klitgord N, Simon C, Boxem M, Milstein S, Rosenberg J, Goldberg DS, Zhang LV, Wong SL, Franklin G, Li S, Albala JS, Lim J, Fraughton C, Llamosas E, Cevik S, Bex C, Lamesch P, Sikorski RS, Vandenhaute J, Zoghbi HY, Smolyar A, Bosak S, Sequerra R, Doucette-Stamm L, Cusick ME, Hill DE, Roth FP, Vidal M (October 2005). "Towards a proteome-scale map of the human protein-protein interaction network". Nature. 437 (7062): 1173–8. doi:10.1038/nature04209. PMID 16189514. S2CID 4427026.
- Ciana P, Fumagalli M, Trincavelli ML, Verderio C, Rosa P, Lecca D, Ferrario S, Parravicini C, Capra V, Gelosa P, Guerrini U, Belcredito S, Cimino M, Sironi L, Tremoli E, Rovati GE, Martini C, Abbracchio MP (October 2006). "The orphan receptor GPR17 identified as a new dual uracil nucleotides/cysteinyl-leukotrienes receptor". The EMBO Journal. 25 (19): 4615–27. doi:10.1038/sj.emboj.7601341. PMC 1589991. PMID 16990797.