μ-opioid receptor
The μ-opioid receptors (MOR) are a class of opioid receptors with a high affinity for enkephalins and beta-endorphin, but a low affinity for dynorphins. They are also referred to as μ(mu)-opioid peptide (MOP) receptors. The prototypical μ-opioid receptor agonist is morphine, the primary psychoactive alkaloid in opium. It is an inhibitory G-protein coupled receptor that activates the Gi alpha subunit, inhibiting adenylate cyclase activity, lowering cAMP levels.


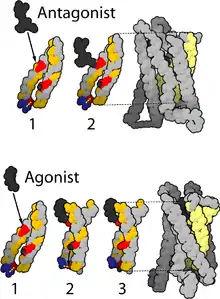
Structure
A cryo-electron microscopy structure of the µ-opioid receptor–Gi protein complex was published in 2018.[6]
Splice variants
Three variants of the μ-opioid receptor are well characterized, though RT-PCR has identified up to 10 total splice variants in humans.[7][8][9]
| μ1 | More is known about the μ1 opioid receptor than the other variants. |
| μ2 | TRIMU 5 is a selective agonist of the μ2 receptor.[10] |
| μ3 | The μ3 variant was first described in 2003.[11] It is responsive to opiate alkaloids but not opioid peptides.[12] |
Location
They can exist either presynaptically or postsynaptically depending upon cell types.
The μ-opioid receptors exist mostly presynaptically in the periaqueductal gray region, and in the superficial dorsal horn of the spinal cord (specifically the substantia gelatinosa of Rolando). Other areas where they have been located include the external plexiform layer of the olfactory bulb, the nucleus accumbens, in several layers of the cerebral cortex, and in some of the nuclei of the amygdala, as well as the nucleus of the solitary tract.
Some MORs are also found in the intestinal tract. Activation of these receptors inhibits peristaltic action which causes constipation, a major side effect of μ agonists.[13]
Activation
MOR can mediate acute changes in neuronal excitability via suppression of presynaptic release of GABA. Activation of the MOR leads to different effects on dendritic spines depending upon the agonist, and may be an example of functional selectivity at the μ-receptor.[14] The physiological and pathological roles of these two distinct mechanisms remain to be clarified. Perhaps, both might be involved in opioid addiction and opioid-induced deficits in cognition.
Activation of the μ-opioid receptor by an agonist such as morphine causes analgesia, sedation, slightly reduced blood pressure, itching, nausea, euphoria, decreased respiration, miosis (constricted pupils), and decreased bowel motility often leading to constipation. Some of these effects, such as analgesia, sedation, euphoria, itching and decreased respiration, tend to lessen with continued use as tolerance develops. Miosis and reduced bowel motility tend to persist; little tolerance develops to these effects.
The canonical MOR1 isoform is responsible for morphine-induced analgesia, whereas the alternatively spliced MOR1D isoform (through heterodimerization with the gastrin-releasing peptide receptor) is required for morphine-induced itching.[15]
Deactivation
As with other G protein-coupled receptors, signalling by the μ-opioid receptor is terminated through several different mechanisms, which are upregulated with chronic use, leading to rapid tachyphylaxis.[16] The most important regulatory proteins for the MOR are the β-arrestins arrestin beta 1 and arrestin beta 2,[17][18][19] and the RGS proteins RGS4, RGS9-2, RGS14, and RGSZ2.[20][21]
Long-term or high-dose use of opioids may also lead to additional mechanisms of tolerance becoming involved. This includes downregulation of MOR gene expression, so the number of receptors presented on the cell surface is actually reduced, as opposed to the more short-term desensitisation induced by β-arrestins or RGS proteins.[22][23][24] Another long-term adaptation to opioid use can be upregulation of glutamate and other pathways in the brain which can exert an opioid-opposing effect, so reduce the effects of opioid drugs by altering downstream pathways, regardless of MOR activation.[25][26]
Tolerance and overdoses
Fatal opioid overdose typically occurs due to bradypnea, hypoxemia, and decreased cardiac output (hypotension occurs due to vasodilation, & bradycardia further contributes to decreased cardiac output).[27][28][29] A potentiation effect occurs when opioids are combined with ethanol, benzodiazepines, or barbiturates, which results in an increased risk for overdose to occur.[27][28] Substantial tolerance to respiratory depression develops quickly, and tolerant individuals can withstand larger doses.[30] However, tolerance to respiratory depression is lost just as quickly during withdrawal.[30] Many overdoses occur in people who misuse their medication after being in withdrawal long enough to lose the tolerance to respiratory depression. Less commonly, massive overdoses have been known to cause circulatory collapse.
Opioid overdoses can be rapidly reversed through the use of opioid antagonists, naloxone being the most widely used example.[27]
See also
References
- GRCh38: Ensembl release 89: ENSG00000112038 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000000766 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Zhorov BS, Ananthanarayanan VS (March 2000). "Homology models of mu-opioid receptor with organic and inorganic cations at conserved aspartates in the second and third transmembrane domains". Arch. Biochem. Biophys. 375 (1): 31–49. doi:10.1006/abbi.1999.1529. PMID 10683246.
- Koehl A, Hu H, Maeda S, et al. (2018). "Structure of the µ-opioid receptor-Gi protein complex". Nature. 558 (7711): 547–552. Bibcode:2018Natur.558..547K. doi:10.1038/s41586-018-0219-7. PMC 6317904. PMID 29899455.
- Dortch-Carnes J, Russell K (2007). "Morphine-stimulated nitric oxide release in rabbit aqueous humor". Exp. Eye Res. 84 (1): 185–90. doi:10.1016/j.exer.2006.09.014. PMC 1766947. PMID 17094965.
- Pan L, Xu J, Yu R, Xu MM, Pan YX, Pasternak GW (2005). "Identification and characterization of six new alternatively spliced variants of the human mu opioid receptor gene, Oprm". Neuroscience. 133 (1): 209–20. doi:10.1016/j.neuroscience.2004.12.033. PMID 15893644. S2CID 22410194.
- Xu J, Lu Z, Narayan A, et al. (2017). "Alternatively spliced mu opioid receptor C termini impact the diverse actions of morphine". J. Clin. Invest. 127 (4): 1561–1573. doi:10.1172/JCI88760. PMC 5373896. PMID 28319053.
- Eisenberg RM (1994). "TRIMU-5, a μ2-opioid receptor agonist, stimulates the hypothalamo-pituitary-adrenal axis". Pharmacol. Biochem. Behav. 47 (4): 943–6. doi:10.1016/0091-3057(94)90300-X. PMID 8029266. S2CID 54354971.
- Cadet P, Mantione KJ, Stefano GB (2003). "Molecular identification and functional expression of μ3, a novel alternatively spliced variant of the human μ opiate receptor gene". J. Immunol. 170 (10): 5118–23. doi:10.4049/jimmunol.170.10.5118. PMID 12734358.
- Stefano GB (2004). "Endogenous morphine: a role in wellness medicine". Med. Sci. Monit. 10 (6): ED5. PMID 15173675.
- Chen, Wency (2012). "Opiate-induced constipation related to activation of small intestine opioid μ2-receptors". World Journal of Gastroenterology. 18 (12): 1391–6. doi:10.3748/wjg.v18.i12.1391. PMC 3319967. PMID 22493554.
- Liao D, Lin H, Law PY, Loh HH (February 2005). "Mu-opioid receptors modulate the stability of dendritic spines". Proc. Natl. Acad. Sci. U.S.A. 102 (5): 1725–30. Bibcode:2005PNAS..102.1725L. doi:10.1073/pnas.0406797102. JSTOR 3374498. PMC 545084. PMID 15659552.
- Liu XY, Liu ZC, Sun YG, Ross M, Kim S, Tsai FF, Li QF, Jeffry J, Kim JY, Loh HH, Chen ZF (2011). "Unidirectional Cross-Activation of GRPR by MOR1D Uncouples Itch and Analgesia Induced by Opioids". Cell. 147 (2): 447–458. doi:10.1016/j.cell.2011.08.043. PMC 3197217. PMID 22000021. Lay summary – Washington University in St. Louis Press Release.
- Martini L, Whistler JL (October 2007). "The role of mu opioid receptor desensitization and endocytosis in morphine tolerance and dependence". Current Opinion in Neurobiology. 17 (5): 556–64. doi:10.1016/j.conb.2007.10.004. PMID 18068348. S2CID 29491629.
- Zuo Z (September 2005). "The role of opioid receptor internalization and beta-arrestins in the development of opioid tolerance". Anesthesia and Analgesia. 101 (3): 728–34, table of contents. doi:10.1213/01.ANE.0000160588.32007.AD. PMID 16115983.
- Marie N, Aguila B, Allouche S (November 2006). "Tracking the opioid receptors on the way of desensitization". Cellular Signalling. 18 (11): 1815–33. doi:10.1016/j.cellsig.2006.03.015. PMID 16750901.
- DuPen A, Shen D, Ersek M (September 2007). "Mechanisms of opioid-induced tolerance and hyperalgesia". Pain Management Nursing. 8 (3): 113–21. doi:10.1016/j.pmn.2007.02.004. PMID 17723928.
- Garzón J, Rodríguez-Muñoz M, Sánchez-Blázquez P (May 2005). "Morphine alters the selective association between mu-opioid receptors and specific RGS proteins in mouse periaqueductal gray matter". Neuropharmacology. 48 (6): 853–68. doi:10.1016/j.neuropharm.2005.01.004. PMID 15829256. S2CID 23797166.
- Hooks SB, Martemyanov K, Zachariou V (January 2008). "A role of RGS proteins in drug addiction". Biochemical Pharmacology. 75 (1): 76–84. doi:10.1016/j.bcp.2007.07.045. PMID 17880927.
- Sirohi S, Dighe SV, Walker EA, Yoburn BC (November 2008). "The analgesic efficacy of fentanyl: relationship to tolerance and mu-opioid receptor regulation". Pharmacology Biochemistry and Behavior. 91 (1): 115–20. doi:10.1016/j.pbb.2008.06.019. PMC 2597555. PMID 18640146.
- Lopez-Gimenez JF, Vilaró MT, Milligan G (November 2008). "Morphine desensitization, internalization, and down-regulation of the mu opioid receptor is facilitated by serotonin 5-hydroxytryptamine2A receptor coactivation". Molecular Pharmacology. 74 (5): 1278–91. doi:10.1124/mol.108.048272. PMID 18703670. S2CID 6310244.
- Kraus J (2009). "Regulation of mu-opioid receptors by cytokines". Frontiers in Bioscience. 1: 164–70. doi:10.2741/e16. PMID 19482692.
- García-Fuster MJ, Ramos-Miguel A, Rivero G, La Harpe R, Meana JJ, García-Sevilla JA (November 2008). "Regulation of the extrinsic and intrinsic apoptotic pathways in the prefrontal cortex of short- and long-term human opiate abusers". Neuroscience. 157 (1): 105–19. doi:10.1016/j.neuroscience.2008.09.002. PMID 18834930. S2CID 9022097.
- Ueda H, Ueda M (2009). "Mechanisms underlying morphine analgesic tolerance and dependence". Frontiers in Bioscience. 14: 5260–72. doi:10.2741/3596. PMID 19482614.
- Blok (2017). "Opioid toxicity" (PDF). Clinical Key. Elsevier.
- Hughes CG, McGrane S, Pandharipande PP (2012). "Sedation in the intensive care setting". Clinical Pharmacology. 4 (53): 53–63. doi:10.2147/CPAA.S26582. PMC 3508653. PMID 23204873.
- Passchier J, Goudswaard P, Orlebeke JF, Verhage F (2011). "Migraine and defense mechanisms: psychophysiological relationships in young females". Social Science & Medicine. 26 (3): 343–50. doi:10.1016/0277-9536(88)90399-1. PMC 3347855. PMID 22577457.
- Zhilenko VN, Khoroshilova NV, Efremova VM (December 1989). "[Hygienic evaluation of atactic polypropylene]". Gigiena I Sanitariia. 10 (12): 86–7. doi:10.1208/s12248-008-9056-1. PMC 2628209. PMID 18989788.
External links
- "Opioid Receptors: μ". IUPHAR Database of Receptors and Ion Channels. International Union of Basic and Clinical Pharmacology.
- mu+Opioid+Receptor at the US National Library of Medicine Medical Subject Headings (MeSH)
- Human OPRM1 genome location and OPRM1 gene details page in the UCSC Genome Browser.