IRAK4
IRAK-4 (interleukin-1 receptor-associated kinase 4), in the IRAK family, is a protein kinase involved in signaling innate immune responses from Toll-like receptors. It also supports signaling from T-cell receptors. IRAK4 contains domain structures which are similar to those of IRAK1, IRAK2, IRAKM and Pelle. IRAK4 is unique compared to IRAK1, IRAK2 and IRAKM in that it functions upstream of the other IRAKs, but is more similar to Pelle in this trait. IRAK4 is important for its clinical applications.




Animals without IRAK-4 are more susceptible to viruses and bacteria but completely resistant to LPS challenge.
History
The first IL-1 receptor-associated kinase (IRAK) was observed in 1994 through experiments with murine T helper cell lines D10N and EL-4.[5] Two years later the first experimental member of this family of kinases, IRAK1, was cloned.[6] In 2002, through database searches at the National Center for Biotechnology Information in an attempt to recognize novel members of the IRAK family, a human cDNA sequence which encoded a peptide sharing significant homology with IRAK1 was identified. This cDNA sequence was found to have five amino acid substitutions compared to IRAK1 and was termed IRAK4.[7]
IRAK4 was proposed to be the mammalian homolog of the Pelle gene found in Drosophila melanogaster and was proposed to require its kinase activity in order for it to function in activating NF-κB. It was also proposed by Li et al. that it might function upstream of other IRAKs and possibly cause a cascade of phosphorylation events through its function as an IRAK1 kinase.[7] This idea of a cascade of phosphorylation events was supported by a study where an IRAK4 knockout in mice showed a more severe phenotype than other IRAK knockout experiments and signalling through Toll/IL-1 receptor (TIR) is virtually eliminated.[7]
In 2007 it was found that IRAK4 activity was necessary for activating signal pathways which lead to mitogen-activated protein kinases (MAPK), or Toll-like receptor-mediated immune responses (TLR), but was not essential to T-cell Receptor (TCR) signalling as was originally proposed.[8]
In recent years, the role of IRAK4 in regards to melanoma and other cancers has been investigated. IRAK4 was found to be in higher levels in some lines of melanoma. By reducing the IRAK4 activity it may be possible to identify new chemotherapeutic agents to treat patients with advanced melanoma for which there is presently no effective treatment or cure.[9]
Protein structure
IRAK4 is a threonine/serine protein kinase made up of 460 amino acids, which contains both a kinase domain and a death domain.[7] Its kinase domain exhibits the typical bilobed structure of kinases, with the N-terminal lobe consisting of a five-stranded antiparallel beta-sheet and one alpha helix. The C-terminal lobe is composed mainly of a number of alpha helices.[10] Also contained within IRAK4's N-terminal is an extension of twenty amino acids, which is unique to IRAK4 among kinases, even within the IRAK family.[11] Situated where the two lobes meet is an ATP binding site, which is covered by a tyrosine gatekeeper. Tyrosine as a gatekeeper is believed to be unique to the IRAK family of kinases.[10] The protein also contains three auto-phosphorylation sites, each of which when mutated results in a decrease in the kinase activity of IRAK4.[12]
A structure of the autophosphorylation of the activation loop has been determined in which the activation loop Thr345 of one monomer is sitting in the active site of another monomer in the crystal (PDB: 4U9A, 4U97).[13][14]
Function, mechanism, signalling pathway
Members of interleukin-1 receptor (Il-1R) and the Toll-like receptor superfamily share an intracytoplasmic Toll-IL-1 receptor (TLR) domain, which mediates recruitment of the interleukin-1 receptor-associated kinase (IRAK) complex via TIR-containing adapter molecules. The TIR-IRAK signaling pathway appears to be crucial for protective immunity against specific bacteria but is redundant against most other microorganisms.[15] IRAK4 is considered the “master IRAK” in the mammalian IRAK family because it is the only component in the IL-1/TLR signalling pathway that is absolutely crucial to its functioning. When one of these pathways is stimulated, the cell is triggered to release proinflammatory signals and to trigger innate immune actions. The loss of IRAK4, or its intrinsic kinase activity, can entirely stop signalling through these pathways.[16]
IRAK4 is involved in signal transduction pathways stimulated by the cellular receptors belonging to the Toll/Interleukin-1 receptor superfamily. The Toll-Like Receptors (TLRs) are stimulated by recognition of pathogen-associated molecular patterns (PAMPS), whereas members of the IL-1R family are stimulated by cytokines.[17] Both play an essential role in the immune response. The ligand binding causes conformational changes to the intracellular domain which allows for the recruitment of scaffolding proteins. One of these proteins, MyD88, uses its death domains to recruit, orient, and activate IRAK4. IRAK2 can then be phosphorylated and joins with IRAK4 and MyD88 to form the myddosome complex, which further phosphorylates and recruits IRAK1.[18] The myddosome complex and IRAK1 recruit and activate TNF receptor-associated factor 6 (TRAF6), a ubiquitin protein ligase.[7] TRAF6 can polyubiquitinate IKK-γ as well as itself, which recruits TGF-β activated kinase 1 (TAK1) in order to activate its ability to phosphorylate IKK-β. These pathways both work to degrade IKKγ, which releases NFκB and free it for translocation into the nucleus. Additionally, TAK1 can activate JNK to induce a MAP kinase pathway which leads to AP-1-induced gene expression.[8] Together, AP-1 and NFκB lead to increased cytokine transcription, adhesion molecule production, and release of second messengers of infection.[18]
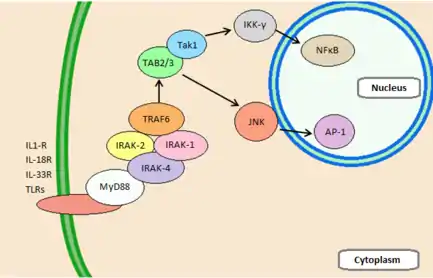
Central to all of these signalling pathways is the kinase IRAK4. Results show that IRAK4 is a crucial component in an animal's response to IL-1. Animals deficient in this kinase were found to be lacking in the ability to recognize viral and bacterial invaders, and were completely resistant to lethal doses of lipopolysaccharide (LPS).[17] This is due to IRAK4's function as both a structural protein and as a kinase. Both of these functions are required for the myddosome complex formation. Additionally, IRAK4 has been shown to be absolutely essential in a TLR signalling. IRAK4 deficient mice have a profoundly impaired ability to produce IL-6, TNF-α, and IL-12 in response to TLR ligands. However it is worthy of note that despite its importance to many immune signalling pathways, IRAK4 does not appear to be involved in TCR signalling.[8]
Clinical significance
There are three components of evidence that illustrate IRAK4's involvement in TLR signalling. First, IRAK4 is the initial kinase near the TLR receptor to activate downstream effectors such as cytokines and chemokines in the inflammatory cascade.[7] Second, deletion of the IRAK4 gene results in various cytokine response defects and finally, patients with IRAK4 deficiency have displayed defective immunity in response to IL-1, IL-8 and other TLR binding ligands.[17] Considering IRAK4's downstream position of these signalling events, it is an important drug therapy target for various inflammatory disorders including rheumatoid arthritis, inflammatory bowel disease and other autoimmune diseases.[18]
Prostate cancer
An important area of research currently being explored is the role the IRAK4 gene may play in the development of prostate cancer. There are several interacting factors that lead to the development of this disease however genetic susceptibility of chronic inflammation has been deemed one of the most important. It has been found that mutations in the IRAK4 gene can lead to dysfunctional TLR signalling and ultimately result in increased innate immune responses and therefore an increased inflammatory response. Over time, this can lead to the onset of prostate cancer.[19]
Melanoma
Another interesting application of the IRAK4 gene was found in a study involving human melanoma patients. This research found that patients with melanin-cell tumors displayed an increase in the phosphorylation state of IRAK4. The siRNA inhibition of IRAK4 in mice displayed greater programmed cell death (PCD) and slowed tumor growth.[18] This experimental study displays yet again another avenue of IRAK4 targeting for therapeutic purposes.
IRAK4 as a drug target
A common concern with IRAK4 drug therapy or knockdown is if its absence would result in unbearable side effects considering IRAK4 plays an extremely central role in the TLR signalling pathway.[16] Children with IRAK4 deficiency have been found to have decreased immunity to some specific bacterial infections yet not to viral, parasitic or other microbe infections. However, as these children enter adulthood and maternal antibodies are no longer present, susceptibility to infections becomes a rarity. In one study, no significant bacterial infections were documented in all investigated patients over the age of 14 with IRAK4 deficiency. This may mean that in later stages of life, IRAK4 inhibition could provide benefits against certain diseases while maintaining immunity.[20]
The next step in this area of research is the formation of safe IRAK4 inhibitors. There has been modest progress in the development of some potential inhibitors of IRAK4 in which their mechanism works by blocking its tyrosine gated ATP binding site. As of 2007 All potential drugs are in the early preclinical stages of development.[21]
Early-stage clinical trials of an IRAK4 inhibitor had started by 2019.[22]
References
- GRCh38: Ensembl release 89: ENSG00000198001 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000059883 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Martin M, Böl GF, Eriksson A, Resch K, Brigelius-Flohé R (July 1994). "Interleukin-1-induced activation of a protein kinase co-precipitating with the type I interleukin-1 receptor in T cells". European Journal of Immunology. 24 (7): 1566–71. doi:10.1002/eji.1830240717. PMID 8026518.
- Cao Z, Henzel WJ, Gao X (February 1996). "IRAK: a kinase associated with the interleukin-1 receptor". Science. 271 (5252): 1128–31. Bibcode:1996Sci...271.1128C. doi:10.1126/science.271.5252.1128. PMID 8599092. S2CID 42977425.
- Li S, Strelow A, Fontana EJ, Wesche H (April 2002). "IRAK-4: a novel member of the IRAK family with the properties of an IRAK-kinase". Proceedings of the National Academy of Sciences of the United States of America. 99 (8): 5567–72. Bibcode:2002PNAS...99.5567L. doi:10.1073/pnas.082100399. PMC 122810. PMID 11960013.
- Kawagoe T, Sato S, Jung A, Yamamoto M, Matsui K, Kato H, et al. (May 2007). "Essential role of IRAK-4 protein and its kinase activity in Toll-like receptor-mediated immune responses but not in TCR signaling". The Journal of Experimental Medicine. 204 (5): 1013–24. doi:10.1084/jem.20061523. PMC 2118579. PMID 17485511.
- Srivastava R, Geng D, Liu Y, Zheng L, Li Z, Joseph MA, et al. (December 2012). "Augmentation of therapeutic responses in melanoma by inhibition of IRAK-1,-4". Cancer Research. 72 (23): 6209–16. doi:10.1158/0008-5472.CAN-12-0337. PMC 3677596. PMID 23041547.
- Wang Z, Liu J, Sudom A, Ayres M, Li S, Wesche H, et al. (December 2006). "Crystal structures of IRAK-4 kinase in complex with inhibitors: a serine/threonine kinase with tyrosine as a gatekeeper". Structure. 14 (12): 1835–44. doi:10.1016/j.str.2006.11.001. PMID 17161373.
- Kuglstatter A, Villaseñor AG, Shaw D, Lee SW, Tsing S, Niu L, et al. (March 2007). "Cutting Edge: IL-1 receptor-associated kinase 4 structures reveal novel features and multiple conformations". Journal of Immunology. 178 (5): 2641–5. doi:10.4049/jimmunol.178.5.2641. PMID 17312103.
- Cheng H, Addona T, Keshishian H, Dahlstrand E, Lu C, Dorsch M, et al. (January 2007). "Regulation of IRAK-4 kinase activity via autophosphorylation within its activation loop". Biochemical and Biophysical Research Communications. 352 (3): 609–16. doi:10.1016/j.bbrc.2006.11.068. PMID 17141195.
- Ferrao R, Zhou H, Shan Y, Liu Q, Li Q, Shaw DE, et al. (September 2014). "IRAK4 dimerization and trans-autophosphorylation are induced by Myddosome assembly". Molecular Cell. 55 (6): 891–903. doi:10.1016/j.molcel.2014.08.006. PMC 4169746. PMID 25201411.
- Xu Q, Malecka KL, Fink L, Jordan EJ, Duffy E, Kolander S, et al. (December 2015). "Identifying three-dimensional structures of autophosphorylation complexes in crystals of protein kinases". Science Signaling. 8 (405): rs13. doi:10.1126/scisignal.aaa6711. PMC 4766099. PMID 26628682.
- Ku CL, Yang K, Bustamante J, Puel A, von Bernuth H, Santos OF, et al. (February 2005). "Inherited disorders of human Toll-like receptor signaling: immunological implications". Immunological Reviews. 203: 10–20. doi:10.1111/j.0105-2896.2005.00235.x. PMID 15661018. S2CID 21786295.
- Wang Z, Wesche H, Stevens T, Walker N, Yeh WC (1 January 2009). "IRAK-4 inhibitors for inflammation". Current Topics in Medicinal Chemistry. 9 (8): 724–37. doi:10.2174/156802609789044407. PMC 3182414. PMID 19689377.
- Suzuki N, Suzuki S, Duncan GS, Millar DG, Wada T, Mirtsos C, et al. (April 2002). "Severe impairment of interleukin-1 and Toll-like receptor signalling in mice lacking IRAK-4". Nature. 416 (6882): 750–6. Bibcode:2002Natur.416..750S. doi:10.1038/nature736. PMID 11923871. S2CID 4428621.
- Chaudhary D, Robinson S, Romero DL (January 2015). "Recent advances in the discovery of small molecule inhibitors of interleukin-1 receptor-associated kinase 4 (IRAK4) as a therapeutic target for inflammation and oncology disorders". Journal of Medicinal Chemistry. 58 (1): 96–110. doi:10.1021/jm5016044. PMID 25479567.
- Sun J, Wiklund F, Hsu FC, Bälter K, Zheng SL, Johansson JE, et al. (March 2006). "Interactions of sequence variants in interleukin-1 receptor-associated kinase4 and the toll-like receptor 6-1-10 gene cluster increase prostate cancer risk". Cancer Epidemiology, Biomarkers & Prevention. 15 (3): 480–5. doi:10.1158/1055-9965.EPI-05-0645. PMID 16537705.
- Picard C, Puel A, Bonnet M, Ku CL, Bustamante J, Yang K, et al. (March 2003). "Pyogenic bacterial infections in humans with IRAK-4 deficiency". Science. 299 (5615): 2076–9. Bibcode:2003Sci...299.2076P. doi:10.1126/science.1081902. PMID 12637671. S2CID 22438404.
- Ku CL, von Bernuth H, Picard C, Zhang SY, Chang HH, Yang K, et al. (October 2007). "Selective predisposition to bacterial infections in IRAK-4-deficient children: IRAK-4-dependent TLRs are otherwise redundant in protective immunity". The Journal of Experimental Medicine. 204 (10): 2407–22. doi:10.1084/jem.20070628. PMC 2118442. PMID 17893200.
- Curis, Inc. (2019). "Curis Provides First-Ever Demonstration that Targeting IRAK4 in Patients with Relapsed/Refractory Non-Hodgkin's Lymphoma Results in Anti-Cancer Activity in Ongoing Phase 1 Study". PR Newswire Association LLC.
External links
- IRAK4+protein,+human at the US National Library of Medicine Medical Subject Headings (MeSH)
- Overview of all the structural information available in the PDB for UniProt: Q9NWZ3 (Human Interleukin-1 receptor-associated kinase 4) at the PDBe-KB.
- Overview of all the structural information available in the PDB for UniProt: Q8R4K2 (Mouse Interleukin-1 receptor-associated kinase 4) at the PDBe-KB.
